Бфл вк: Чемпионат BFL 20/21, BFL. Официальный сайт
Содержание
Что означает BFL? -определения BFL
Вы ищете значения BFL? На следующем изображении вы можете увидеть основные определения BFL. При желании вы также можете загрузить файл изображения для печати или поделиться им со своим другом через Facebook, Twitter, Pinterest, Google и т. Д. Чтобы увидеть все значения BFL, пожалуйста, прокрутите вниз. Полный список определений приведен в таблице ниже в алфавитном порядке.
Основные значения BFL
На следующем изображении представлены наиболее часто используемые значения BFL. Вы можете записать файл изображения в формате PNG для автономного использования или отправить его своим друзьям по электронной почте.Если вы являетесь веб-мастером некоммерческого веб-сайта, пожалуйста, не стесняйтесь публиковать изображение определений BFL на вашем веб-сайте.
Все определения BFL
Как упомянуто выше, вы увидите все значения BFL в следующей таблице. Пожалуйста, знайте, что все определения перечислены в алфавитном порядке.
Вы можете щелкнуть ссылки справа, чтобы увидеть подробную информацию о каждом определении, включая определения на английском и вашем местном языке.
Что означает BFL в тексте
В общем, BFL является аббревиатурой или аббревиатурой, которая определяется простым языком. Эта страница иллюстрирует, как BFL используется в обмена сообщениями и чат-форумах, в дополнение к социальным сетям, таким как VK, Instagram, Whatsapp и Snapchat. Из приведенной выше таблицы, вы можете просмотреть все значения BFL: некоторые из них образовательные термины, другие медицинские термины, и даже компьютерные термины. Если вы знаете другое определение BFL, пожалуйста, свяжитесь с нами. Мы включим его во время следующего обновления нашей базы данных. Пожалуйста, имейте в информации, что некоторые из наших сокращений и их определения создаются нашими посетителями. Поэтому ваше предложение о новых аббревиатур приветствуется! В качестве возврата мы перевели аббревиатуру BFL на испанский, французский, китайский, португальский, русский и т.
д. Далее можно прокрутить вниз и щелкнуть в меню языка, чтобы найти значения BFL на других 42 языках.
BlackPrince 11:46 ЭЦЛКЩВШЫИКИКТК DoctorWow 11:46 ДА СУКА ВЫ ЗАЕБЛИ! YellowApple 11:47 ЭТО ТЫ ЗАЕБ, МЫ ИЕШАЕШЬ НАМ СРАТЬСЯ!!! SuperSquirrel 11:47 ПАШОЛ НАХУЙ YellowApple 11:47 БЛЯЯЯЯЯЯЯЯТЬ won9erlan9 11:48 ААААААААААААААААА, СОНХВА РУГАЕТСЯ!!!!!! IllusionYesNo 11:48 ЮНХО, ЗАТКНИСЬ!!! won9erlan9 11:48 Что происходит, _, YourBoyfriend 11:49 Пхпхпхпх, мы мешаешь жылчлытвлв Jeong-in_F-k 11:50 Слепой баран Jeong-in_F-k 11:50 Пиздец. … Seungminmua11
11:51 Сынмин-и, тихо тихо… Daddy666 11:51 Сохо, ты слишком недоразвит, чтобы разбрызгивать своими слюнями на такого бога, как я SuperSquirrel 11:52 А ты слишком ничтожество, чтобы даже с таким недоразвитым, как я, вести диструктивный диалог, иди в пизду Sexy_Soul 11:53 *всё ещё пытается вспомнить как выглядит сколопендра*🤔🤔 IllusionYesNo 11:54 Дылвлкк Jeong-in_F-k 11:55 ОБОЖАБ ЭТОТ ХАОС! Jeong-in_F-k 11:55 Я готов даже дать Джисону и Сохо, лишь бы они продолжали эту поебень! Jeong-in_F-k 11:56 Я не педафил… Sexy_Soul 11:56 Я тоже
SuperSquirrel 11:57 На говно похоже) Sexy_Soul 11:57 😝 SuperSquirrel 11:57 🖕✍️ Sexy_Soul 11:57 Шо SuperSquirrel 11:57 Блять Sexy_Soul 11:58 Ай, да пожуй Sexy_Soul 11:58 Дуьктулыьу Bfl. baby 11:58 Ну вы и долбаёбы) Bfl.baby 11:59 Продолжайте, я уже взял попкорн Jeong-in_F-k 11:59 А я курочку Bfl.baby 12:00 Вы 0хуели Sexy_Soul 12:00 Джисооооооооооон Oh_my.Blblblblbl 12:00 Чо SuperSquirrel
12:01 Поговорить надо, чо
Не чокай тут на меня :/ Oh_my.Blblblblbl 12:02 Лан. SuperSquirrel 12:02 :-:::: ChangbinGrrr 12:03 Почему он не послал? ChangbinGrrr 12:03 Ну Чанбини, ну ты конечно тугодум Jeong-in_F-k 12:06 :0 Daddy666 12:07 … Jeong-in_F-k 12:09 Ещё скажи что и ты Jeong-in_F-k 12:10 …….. Daddy666 12:11 РеальннА Solar_boy 12:12
Oh_my.Blblblblbl, SuperSquirrel
Если я скажу, что хочу на свидание, что ты мне скажешь в ответ? Oh_my.Blblblblbl 12:04 Что SuperSquirrel 12:05 Что Oh_my.
Blblblblbl 12:05 Ну что что SuperSquirrel 12:05 Ну что что Oh_my.Blblblblbl 12:05 Что мы с тобой пойдём на свидание SuperSquirrel 12:06 Что… Oh_my.Blblblblbl 12:07 Эцьаькдвькьк Oh_my.Blblblblbl 12:07 ДЖИИИИ Oh_my.Blblblblbl 12:07 *прыг на ручки к Джи* Oh_my.Blblblblbl 12:07 Ты самый самый лучший парень на свете! Oh_my.Blblblblbl 12:08 После Уёна, конечно же) Oh_my.Blblblblbl 12:08 Пхпхпх, даже спорить не буду) SuperSquirrel 12:09 Аооааоаооаао
Ну тк когда? Oh_my.Blblblblbl 12:10 Подожди ещё
У Хёнджина с Фелом тоже свиданка намечается на ближайшие дни, средь недели что-то вроде
А я тоже средь недели выходной взял на работе, ибо давление что-то в последнее время разыгралось
SuperSquirrel 12:13 :»00
Ну
Мы можем перенести, если ты себя плохо чувствуешь т-т Oh_my.Blblblblbl 12:15 Нет, нет, нет, всё нормально)
Таблетку выпью и хоть в горы! SuperSquirrel 12:16 Мммммм)))
В меня, это хорошо)) Oh_my. Blblblblbl 12:16 .. SuperSquirrel 12:17 Боже, Сан SuperSquirrel 12:17 Всё вот ты, да опошлишь :00
SuperSquirrel 12:17 Я просто всегда тебя хочу, что я могу поделать? Oh_my.Blblblblbl 12:18 ,_, SuperSquirrel 12:18 Ты хоть знаешь, как выглядят твои сука руки?? Oh_my.Blblblblbl 12:18 Ты тоже очень красивый и классно сложен… SuperSquirrel 12:19 Не так как ты
Я на тебя слюнки пускаю) Oh_my.Blblblblbl 12:20 ВСЁ, ПОЙДУ ПОГОВОРЮ С ХЁНДЖИНОМ SuperSquirrel 12:21 Иди, свет очей моих ~ ❤️ Oh_my.Blblblblbl 12:21
SuperSquirrel, I’m_sexy_and_I_know_it
Привет SuperSquirrel 12:25 Хааааай I’m_sexy_and_I_know_it 12:29 Какие планы на будущее? SuperSquirrel 12:30 Много каких
А что? I’m_sexy_and_I_know_it 12:31 Думаю над идеей собраться всем вместе в честь выписки Уёна SuperSquirrel 12:32 Ааааа I’m_sexy_and_I_know_it 12:32 Нуууу🤔🤔🤔 I’m_sexy_and_I_know_it 12:32 А сёдня какой день недели вообще? I’m_sexy_and_I_know_it 12:32 Вторник SuperSquirrel 12:32 В четверг у меня свиданка с Фелом I’m_sexy_and_I_know_it 12:33 А так свободен полностью буду только на выходных, в воскресенье, хотя с каких пор это считается выходным днём, пхпэжп I’m_sexy_and_I_know_it 12:34 Ну да SuperSquirrel 12:35 А чо ит ты вдруг занялся этим вопросом, а не друзья Уёна? I’m_sexy_and_I_know_it 12:35 Сан и настоял, чтобы я поговорил со всеми SuperSquirrel 12:36 Ааа I’m_sexy_and_I_know_it 12:37 Ясно тогда I’m_sexy_and_I_know_it 12:37 Заодно подарок подарим I’m_sexy_and_I_know_it 12:37 А
Точно SuperSquirrel 12:40 Пэпжпззпзп I’m_sexy_and_I_know_it 12:41 Ну ты как Чан I’m_sexy_and_I_know_it 12:41 Да бля, бывает жвьатв SuperSquirrel 12:41 Ладно, спасибо SuperSquirrel 12:41 Не за что I’m_sexy_and_I_know_it 12:41 Пожалуйста I’m_sexy_and_I_know_it 12:41 Пхпхпх SuperSquirrel 12:42
SuperSquirrel, Oh_my. Blblblblbl
Кароче в пятницу SuperSquirrel 12:44 Оаоаоа, хорошо~ Oh_my.Blblblblbl 12:47
won9erlan9, Bfl.baby
Ёсан-и, привет
есть какие-то варианты поздравлений в честь выписки Уёна? won9erlan9 15:56 Привет, хён~
Не знаю, я хочу отдельно с Уёном посидеть, чтобы без кого-либо
Мне много что ему надо рассказать — Bfl.baby 16:01 Ааа, хорошо won9erlan9 16:02 Да и вообще я соскучился
Я не мог даже чаще навещать его. Мне до сих пор кажется, что я виноват. Bfl.baby 16:03 Ты не виноват, успокойся (
Всё хорошо! won9erlan9 16:04 Да, наверное.. Bfl.baby 16:05 Так, не распускать нюни! won9erlan9 16:05 Хочешь, можешь сегодня ко мне приехать? won9erlan9 16:06 Хочу. Bfl.baby 16:07 ~~ won9erlan9 16:08 Тогда приготовлю что-нибудь
Что ты хочешь? won9erlan9 16:08 Много чего Bfl. ~ won9erlan9 16:14
Зоопарк
Мне вот интересно DoctorWow 19:02 Где Уён и Хонджун? IllusionYesNo 19:07 Угу DoctorWow 19:08 Взылты BlackPrince 19:09 У них скорее всего трах марафон #DinDonbitch 19:10 lol ChangbinGrrr 19:11 lol Solar_boy 19:13 lol SuperSquirrel 19:13 lol YourBoyfriend 19
Lütfen aleti çalıştırmadan önce kullanma kılavuzunu dikkatli
biçimde okuyun.
Před spuštěnám stroje si pečlivě pročtěte návod k používání.
Pred prvým použitím prístroja si pozorne prečítajte návod na
obsluhu.
Przed uruchomieniem elektronarzędzia zapoznać się uważnie z
treścią instrukcji.
Kérjük alaposan olvassa el a tájékoztatót mielőtt a gépet
használja.
Prosimo, da pred uporabo pozorno preberete to navodilo za
uporabo.
Molimo da pažljivo pročitate uputu o upotrebi prije puštanja u rad.
Pirms sākt lietot instrumentu, lūdzu, izlasiet lietošanas pamācību.
Prieš pradėdami dirbti su prietaisu, atidžiai perskaitykite jo
naudojimo instrukciją.
Palun lugege enne käikulaskmist kasutamisjuhend hoolikalt läbi.
Пожалуйста, внимательно прочтите инструкцию по
использованию перед началом любых операций с
инструментом.
Преди пускане на уреда в действие моля прочетете
внимателно инструкцията за използване.
Va rugăm citiţi cu atenţie instrucţiunile înainte de pornirea maşinii
Ве молиме внимателно прочитајте ги упатствата прд да ја
вклучите машината.
使用本机器之前请详细阅读使用说明书。
Remove the battery pack before starting
any work on the machine.
Vor allen Arbeiten an der Maschine den
Wechselakku herausnehmen
Avant tous travaux sur la machine retirer
l’accu interchangeable.
Prima di iniziare togliere la batteria dalla macchina.
Retire la batería antes de comenzar cualquier trabajo en
la máquina.
Antes de efectuar qualquer intervenção na máquina
retirar o bloco acumulador.
Voor alle werkzaamheden aan de machine de akku
verwijderen.
Ved arbejde inden i maskinen, bør batteriet tages ud.
Ta ut vekselbatteriet før du arbeider på maskinen
Drag ur batteripaket innan arbete utföres på maskinen.
Tarkista pistotulppa ja verkkojohto mahdollisilta
vauriolta. Viat saa korjata vain alan erikoismies.
Ðñéí áðü êÜèå åñãáóßá óôç ìç÷áíÞ áöáéñåßôå ôçí
áíôáëëáêôéêÞ ìðáôáñßá.
Aletin kendinde bir çalışma yapmadan önce kartuş
aküyü çıkarın.
Před zahájením veškerých prací na vrtacím šroubováku
vyjmout výměnný akumulátor.
Pred každou prácou na stroji výmenný akumulátor
vytiahnuť.
Przed przystąpieniem do jakichkolwiek prac na
elektronarzędziu należy wyjąć wkładkę akumulatorową.
Karbantartás, javítás, tisztítás, stb. előtt az akkumulátort
ki kell venni a készülékből.
Pred deli na stroju izvlecite izmenljivi akumulator.
Prije svih radova na stroju izvaditi bateriju za zamjenu.
Pirms mašīnai veikt jebkāda veida apkopes darbus, ir
jāizņem ārā akumulātors.
Prieš atlikdami bet kokius darbus įrenginyje, išimkite
keičiamą akumuliatorių.
Enne kõiki töid masina kallal võtke vahetatav aku välja.
Bûíüòå àêêóìóëÿòîð èç ìàøèíû ïåðåä
ïðîâåäåíèåì ñ íåé êàêèõ-ëèáî ìàíèïóëÿöèé.
Преди започване на каквито е да е работи по
машината извадете акумулатора.
Scoateţi acumulatorul înainte de a începe orice
intervenţie pe maşină.
Извадете го батерискиот склоп пред отпочнување на
каков и да е зафат врз машината.
在机器上进行任何修护工作之前,先拿出蓄电池。
BFL 18
Original instructions
Originalbetriebsanleitung
Notice originale
Istruzioni originali
Manual original
Manual original
Oorspronkelijke
gebruiksaanwijzing
Original brugsanvisning
Original bruksanvisning
Bruksanvisning i original
Alkuperäiset ohjeet
Πρωτότυπο οδηγιών χρήσης
Orijinal işletme talimatı
Původním návodem k používání
Pôvodný návod na použitie
Instrukcją oryginalną
Eredeti használati utasítás
Izvirna navodila
Originalne pogonske upute
Instrukcijām oriģinālvalodā
Originali instrukcija
Algupärane kasutusjuhend
Оригинальное руководство по
эксплуатации
Оригинално ръководство за
експлоатация
Instrucţiuni de folosire originale
Оригинален прирачник за работа
原始的指南
история развития, ASIC / Habr
Bitcoin-майнер от ASICMINER, производительностью ~2TH/s(источник)
Предыдущая часть.
В этот раз рассмотрим заключительную часть истории развития вычислительных систем, предназначенных для добычи биткоинов, а именно эпоху ASIC, поставившую крест на GPU и FPGA майнерах.
Butterfly Labs (BFL)
BFL были первой компанией анонсировавшей свой майнер на основе ASIC. Уверенность в успехе во многом была основана на предыдущих достижениях этой компании в области FPGA-майнеров.
В июне 2012 BFL начала принимать предзаказы на три типа майнеров: $149 за Jalapenos, производительностью 4.5 GH/s; $1299 за SC Singles при 60GH/s; и за $30K можно было предзаказать монструозный SC MiniRigs с производительностью1500GH/s.
При такой цене эти майнеры могли добывать в 20-50 раз больше биткоинов на доллар инвестиций, чем GPU или FPGA майнеры. Сумма предзаказов только за первый день привысила $250K и активность заказчиков еще долгое время оставалось высокой. Этого оказалось более чем достаточно, чтобы покрыть существенные расходы на проектирование микросхем и изготовление литографических масок для 65нм техпроцесса, которые оцениваются в $500K.
Каждый чип производства BFL содержит 16 полностью развернутых конвейеров для вычисления SHA256, что по производительности эквивалентно 16 mid-hi-end FPGA, умещенным в 1 ASIC. При этом размер чипа составлял всего лишь 10х10мм в BGA корпусе.
Но тут ВНЕЗАПНО!… Первоначально BFL планировала начать поставки майнеров в начале ноября. Однако сроки многократно сдвигались из-за задержек связанных с производством на фабрике, сборкой и проблемами самой BFL. Также, энергопотребление на уровне 0.8W на GH/s за месяц до появления внезапно подскочило до 1.2W, а корпус сменился с QFN на BGA.
Энергоэффективность микросхем в итоге стала их основной проблемой. Энергопотребление оказалось в 4-8 раз выше планируемого, что вынудило снизить тактовую частоту вдвое с 500MHz до 250MHz. Из-за этого также пришлось вносить изменения во всё оборудование, которое проектировалось под эти чипы. Например, майнер Jalapenos изначально должен был использовать единственный ASIC чип, но, для достижения заявленной производительности в 4. 5GH/s, потребовалось использование двух чипов. А итоговое энергопотребление составило ~6W на 1GH/s.
А еще оказалось, что только у 60% функционируют все 16 конвейеров, еще у 20% — 15 конвейеров, у 15% — 14, а оставшиеся 5% могут похвастаться лишь 12-13 работающими конвейерами.
Страсти в интернетах
Совершенно ожидаемо, что, в свете этих событий, на интернет-форумах развернулась нешуточная драма. Заказчики, которые профинансировали компанию миллионами долларов, требовали объяснений задержкам поставок. Разнообразные срывы сроков и уточнение спецификаций привели к тому, что клиенты получили первые образцы оборудования только в апреле 2013го. На пять месяцев позже запланированных сроков и почти год после момента фактической оплаты. Огромная задержка поставок существовала до августа 2013.
Клиенты BFL небезосновательно считали, что их покупки стремительно устаревают, даже еще до того, как они успели получить их в руки. Однако изначальные прогнозы относительно сроков разработки, изготовления и масштабных поставок были весьма оптимистичными с самого начала. Особенно, если учесть, что это первый ASIC продукт для BFL. Фактические сроки оказались вполне типичными для данной индустрии, ведь, например, у Intel уходит около года с появления первых тестовых образцов и до начала поставок.
А вот что совсем нетипично, так это уровень прозрачности, который обеспечивала BFL для своих клиентов на протяжении этого времени. Возможно, это объясняется моделью финансирования, основанной на предзаказах. С другой стороны это привело к росту раздражения и злобы среди клиентов, с тревогой следящих за графиком сложности майнинга BTC и гадающих, не поставили ли они не на ту лошадку…
ASICMINER
Деятельность этой компании началась в июле, после того как BFL начала принимать предзаказы на свои майнеры. Компания была основана тремя гражданами Китая и одной из их мотиваций было создать конкуренцию для BFL и не дать той стать единственным поставщиком майнеров. Подход ASICMINER к ведению дел сильно отличался от использованного BFL, поскольку они не располагали тем доверием, что заработала последняя на поставках FPGA-майнеров.
Также примечательно, что сбор средств был проведен исключительно через интернет форумы. Основатели подробно излагали свои планы по разработке ASIC и отвечали на сотни вопросов от онлайн сообщества, многие из которых касались глубоко технических деталей их бизнес модели технических решений и финансовой благонадежности.
К 18 июля была зарегистрирована компания, юридически расположенная в городе Шенжень на юге Китая, был подписан контракт c производителем микросхем и получены технологические файлы, необходимые для того, чтобы начать проектирование микросхемы. Уже к 29 июля была готова первая версия дизайна, которая должна была обеспечить производительность 1.25GH/s на каждый чип, при техпроцессе 130нм, площади чипа 17.5 мм кв. и энергопотреблении 13.3W. Техпроцесс 130нм был выбран по причине низких капитальных затрат на разработку – приблизительно $150K. Согласно постам, использовался стандартный промышленный маршрут проектирования: Verilog, имитационное моделирование в VCS, синтез на основе Verdi, IC Compiler для трассировки и размещения, Calibre для проверки правил проектирования (DRC) и т. д. Набор лицензий на эти средства стоил бы немалых денег где-нибудь в США или Европе, но в Китае и лицензии, как и рабочая сила, оказались гораздо дешевле.
В начале августа, когда была завершена первая итерация проектирования, размещения и трассировки, компании удалось привлечь средства через IPO, проведенное на онлайн-бирже GLBSE, осуществлявшей торги в bitcoin. Было предложено продавать 1 из 400 тыс. акций компании за 0.1 BTC, при этом акционерам отходило не более 200 тыс. акций. Бизнес план предполагал начать майнинг на своём собственном оборудовании с общей производительностью 12TH/s, а затем начать прямые продажи чипов или оборудования покупателям. Прибыль должна была распределяться между акционерами, пропорционально их доле в компании.
IPO завершилось 27 августа, всего было продано 163.962 акции, что на тот момент примерно равнялось $160К. Интересен тот факт, что голоса акционеров уже на этом этапе использовались для того, чтобы принять решение о конвертации 8000BTC, из собранных в ходе IPO, в привычную валюту, чтобы минимизировать риски связанные с курсом биткоина и необходимостью оплаты услуг фабрики-производителя чипов.
22 сентября была опубликована финальная спецификация чипа: 1.05V, 335MHz, 6×6 мм. и 4.2W на GH/s производительности. Дизайн чипа практически повторял открытый дизайн, используемый ранее для FPGA-майнеров, но с большей частотой, меньшим энергопотреблением и во много раз дешевле.
6го октября биржа GLBSE закрылась из-за взлома и разногласий среди её основателей. Поскольку информация об акционерах хранилась биржей анонимно, представители ASICMINER больше не знали кто их акционеры. Ситуацию усугубил и тот факт, что часть средств, привлеченных в ходе IPO, застряла на счетах биржи. Со временем, на основе электронной почты и других документов, удалось восстановить владельцев примерно 150 тыс. акций. На это ушло два месяца.
К 14 октября был запущен процесс производства и к 31 октября первые вафли добрались до стадии металлизации, за которой следует разрезание вафли на отдельные чипы. Однако фабрика задержала производство, в интересах других, более крупных заказов, и первые готовые чипы появились только 28 декабря.
14 февраля общая вычислительная мощность майнеров, которую ввёл в строй ASICMINER, достигла отметки 2TH/s. В дальнейшем мощность только увеличивалась, но не без всевозможных проблем.
Наконец компания приступила к прямым продажам оборудования. Сначала было продано около 60 «лезвий», чипы которых давали 10.7GH/s @ 83W при цене 50-75 биткоинов (примерно $5K-7.5K). Затем компания разработала майнер в стиле USB-стика под названием Block Erupter. Майнер, содержащий единственный ASIC чип, изначально продавался по 2BTC и стремительно падал в цене. Сейчас его можно найти на Amazon в районе $50-60.
USB-хаб с целым массивом Block Erupter’ов, каждый из которых дает 330 MH/s или примерно половину производительности GPU AMD 7970. (источник)
Avalon
Еще одна компания, которой удалось привлечь инвестиции путем прямых предпродаж через интернет. Её история не столь интересна, поэтому не буду тратить на неё много времени. ASIC выпускаемые этой компанией используют 110нм TSMC-техпроцесс, каждый чип размером 4х4 мм содержит единственный SHA256 конвейер, а типовая поставка представляет собой 300 чипов, смонтированых на поверхности трех «лезвий» в 4U форм-факторе. Как и ASICMINER, компания расположена в Китае.
Первоначально заказы размещались на майнеры, производительностью 66GH/s @ 600W и ценой $1299. Затем появились майнеры в формате USB-stick. А сейчас компания ведет разработку ASIC на технологии 55nm.
Bitcoin Hardware Scaling
На текущий момент многим энтузиастам майнинга удалось предзаказать майнеры, которые будут изготовлены по технологии 28 и менее нанометров. Это вызывает закономерный вопрос – насколько чипы биткоин-майнеров масштабируются в соответствии с техпроцессом. Из-за проблем тёмного кремния, улучшения в производительности следующего поколения микросхем оказываются ограниченными энергоэффективностью на уровне ~1.4x на каждый шаг техпроцесса. Логика биткоин-майнера представляет собой едва ли не худший для тёмного кремния случай, еще более худший чем Multicore или GPU, по причине практически полного отсутствия накристальной памяти и высокой активности всех элементов схемы. Т.е. если, допустим, прогресс остановится на 10нм, то можно ожидать лишь ~6. 5х улучшения производительности, по сравнению с 65нм чипами. История когда BFL пришлось снижать частоту своих чипов из-за ограничений энергопотребления лишь еще раз показывает к чему сейчас приводят попытки вылезти за рамки «дозволенного» тёмным кремнием.
После того, как стремительная смена поколений майнеров перешла к переходу на ASIC, разница между последующими поколениями майнеров будет не настолько велика, чтобы быстро отправить на пенсию старое железо. Полная аналогия с ПК – если в середине 90х рост производительности превращал топовое железо в хлам за 2-3года, то сейчас и 5-летнее железо выглядит очень даже неплохо. Для производителей ASIC это будет означать погоню за энергоэффективностью, а не производительностью. А среди групп энтузиастов майнинга наибольшую выгоду получит тот, у кого будет доступ к наиболее дешевой электроэнергии. Уже сейчас известны случаи, когда скупалось старое оборудование для майнинга и возвращалось в строй, скажем, вблизи электростанций, где тарифы на электроэнергию намного ниже городских.
Оптимизация энергоэффективности
Чипы от BFL по технологии 65нм потребляют ~5.5W на GH/s. Для 130нм-ASICMINER и 110нм-Avalon чипов эти цифры составляют 8W и 9W соответственно. Согласно закону Деннарда, при переходе на 14нм можно ожидать 65/14 = 4.6x снижение энергопотребления, примерно до уровня 1W на GH/s.
Правда, т.к. первые поколения ASIC-майнеров ставили своей целью перейти от FPGA в эпоху ASIC, то, вероятно, есть огромный простор для дальнейших оптимизаций архитектуры и схемотехники с целью сократить энергопотребление, не ухудшив производительность и не увеличивая площадь. Среди таких оптимизаций может оказаться использование нескольких уровней напряжения, замена триггеров на latches или даже использование самосинхронной схемотехники, чтобы сэкономить энергию, которая тратится на тактирование. Согласно экспертным оценкам, есть еще по крайней мере четырехкратная возможность снижения энергопотребления в рамках используемого техпроцесса.
Помимо базовых оптимизаций существует много улучшений, характерных для тёмного кремния, и для которых ASIC-майнер отлично подходит. Например, использование уровней напряжения, близких к порогу срабатывания (NTV – Near Threshold Voltage). Отсутствие статической памяти и необходимости передавать сигналы на большое расстояние внутри чипа позволяет ожидать здесь еще 5х снижения энергопотребления.
Также, поскольку аппаратная реализация SHA256 очень проста, то можно представить специализированный техпроцесс изготовления для майнеров, по аналогии с разными процессами, применяемыми для чипов памяти и логики сегодня.
Вместо заключения
Развитие bitcoin-майнеров привело к появлению специализированных решений на основе ASIC. Без всякой поддержки со стороны крупных компаний. Пользователи самоорганизовались, сами профинансировали разработку программной и аппаратной части, преодолели риски и недоверие, оценили бизнес-планы и справились с задачей создания дорогих чипов при невероятно малом бюджете. Это неслыханное дело в наше время, где оценка усилий по созданию микросхем последнего поколения начинается с отметки $100млн. , а количество специализированных чипов падает год от года.
Какие из этого можно извлечь уроки? При каких условиях создание чипов под конкретную задачу действительно возможно? Несколько мыслей на этот счет.
Чипы, производимые на заказ, наиболее конкурентноспособны по сравнению с массовым производством, если проходят «концентрационный тест» — выгода от использования специализированного чипа должна заключаться в нем самом, а не других компонентах системы. В случае биткоина, прибыльность – прямая характеристики чипа. Лишь с несколькими дополнительными факторами, вроде цен на электричество.
Если вычисления обладают высоким параллелизмом, а специализированная реализация гораздо меньше чем эквивалентный ей код для системы общего назначения, то ASIC – хороший выбор для того, чтобы сделать скачок в производительности.
Как ни странно, но исследования, проводимые в университетах, сыграли весьма скромную роль в разработке ASIC-майнеров. Для этого есть объяснение. Во-первых, университетские исследования направлены на новейшие технологии производства, которые из-за своей нестабильности не подходят для быстрого использования. Во-вторых, у университетов есть бесплатный доступ к САПР, стоимостью десятки миллионов долларов, лицензированным только для некоммерческого использования. Это означает, что выпускник университета просто не знает как делать «железо» дешево, не разорившись на лицензиях.
Время выхода на рынок обратно пропорционально новизне используемого техпроцесса. 130нм появится первым, 65нм появится потом. В тех случаях, где использование ASIC действительно дает выигрыш, нет большой разницы в том, какой техпроцесс используется. Зато очень важно время.
Инвестиционный капитал также не сыграл значительной роли. Во многом благодаря распространенному мнению, что «железные» стартапы слишком дороги и занимают слишком много времени. Пользователи смогли сами профинансировать проект, даже не прибегая и к помощи сайтов вроде kickstarter.
Машины для майнинга биткоинов имеют очень сильное и понятное людям объяснение своей ценности – «вы покупаете майнер и он приносит вам много денег». Кроме того, эта идея уже была проверена на предыдущих поколениях майнеров.
Две из компаний-разработчиков оказались из Китая и там же вели свою деятельность. Хоть Кремниевая Долина и известна тем, что стоит на острие прогресса, большая часть работы в условиях малых бюджетов выполняется в Азии. Дешевый доступ к рабочей силе и лицензиям играет важную роль. Маршрут проектирования, использованный ASICMINER, в США обошелся бы в $400K за лицензии на каждого разработчика.
Для того, чтобы добиться инноваций в железе, необходимо перепробовать множество новых идей, при малых затратах. Но современная полупроводниковая индустрия отошла от этого направления в сторону дорогих и очень дорогих чипов. В результате «полупроводниковые» стартапы практически не существуют, и практически нет областей, где можно было бы исследовать рискованные инновационные идеи. Не потому что нет идей, а потому, что уж очень дорого. В то же время сокращается спрос на hardware-engineer’ов на рынке труда, а молодые специалисты уходят в софтверные компании за более высокими зарплатами. Что-то с этим надо делать.
CC-BFL — Airbus A320-214 — Latam Airlines
23 января 2022
23 Jan 2022
—
23 января 2022
Cordoba (COR)
LIM)
LA2436
—
09:20
—
—
13:10
Play
22 января 2022
—
Ориентировочный вылет 16:55
22 января 2022
Cusco (CUZ)
LIMA (LIM)
LA2014
—
21:55
—
23:15
Оценочная вылет 16 :55
Играть
22 января 2022 г.
—
Оценочный вылет 14:20
22 января 2022
Lima (CUZ)
CUSCO (CUZ)
LA2015
—
9:20
—
—
6 20:35
Оценочная вылет 14:20
Play
3
7
22 января 2022
22 Jan 2022
—
—
0 22 января 2022
Cajamarca (CJA)
Lima (Lim)
LA2297
—
16:15
—
9006 —
7
22 января 2022
—
Оценочный отъезд 09 :00
22 января 2022
Лима (LIM)
Каджам Arca (CJA)
LA2296
—
8 14:00
—
—
2 15:15
5
KML CSV Play
3
21 января 2022
9 CANCUN (CUN)
LIMA (LIM)
LA2457
—
—
—
—
—
—
Play
21 января 2022
5: 02
приземлились 04:21
21 января 2022
Cancun (CUN)
LIMA (LIM)
LA2457
5:02
5:10
23:19
04:30
Приземлился 04:21
KML CSV Играть
21 января 2022
5:07
5:07
приземлились 21:45
21 января 2022
Lima (Cun)
9 Cancun (CUN)
LA2456
5:07
16:35
16:38
22:00
приземлился 21:45
kml CSV Play
20 января 2022
1:31
1:58
20 января 2022
Tumbes (TBP)
LIMA (LIM)
La2335
LA2335
1:31
21:25
21:28
21:05
по суше 22:58
KML CSV Play
6
20 января 2022
1:26
приземлились 20:26
9 20 января 2022 9 0020 Lima (LIM)
(TBP)
LA2334
1:40
18:40
18:00
19:00
3 20:31
приземлились 20:26
kml CSV Play
20 января 2022
1:18
30:42
20 января 2022
Puerto Maldonado (PEM)
Lima (Lim)
LA2572
1:18
16:29
16:24
17:55
1 17:55
постучал 17:42
kml CSV Play
20 января 2022
1:16
20 января 2022
Лима (LIM)
Пуэрто-Мальдонадо (PEM)
LA2573
1:16
14:05
14:12
15:45
15:28
KML CSV Play
0
4 19 Jan 20229 Cancun (Cun)
Lima (Lim)
LA2457
LA2457
—
—
—
—
—
4 Play
93 20 янв.
—
—
—
—
—
4 Unnown
8
19 Jan 2022
4:47
4:34
1 19 Jan 2022
Канкун (CUN)
Лима (LIM)
LA2457
90 004 4:47
22:51
23:47
23:47
04:10
приземлились 04:34
kml CSV Play
6
19 января 2022
5:02
приземлился 21:19
9 19 января 2022
Lima (Cun)
9 Cancun (CUN)
LA2456
5:02
5:10
16:17
21:41
приземлился 21:19
kml CSV Play
19 Jan 2022
Lima (Cun)
9 Cancun (CUN)
LA2456
—
—
—
—
—
Неизвестно
Играть
19 января 2022
Сантьяго (S CL)
Lima (Lim)
LA2478
—
—
—
—
—
Play
2
19 Jan 2022
Lima (LIM)
LA2478
—
—
—
—
—
Play
1
19 Jan 2022
3:28
21
19 Jan 2022
Lima (Lim)
LA2478
LA2478
3:28
4 10:45
10:53
14:40
21
KML CSV Play
19 января 2022 г. 90 007
2:58
2:58
приземлился 08:57
19 Jan 2022 Lima (Lim)
19 Santiago (SCL)
LA2479
2:58
2:58
05:58
09:15
kml CSV Play
9004
19 Jan 2022 Lima (Lim)9 Сантьяго (SCL)
LA2479
—
—
—
—
—
Unnown
3
90040 18 января 20229 Cancun (Cun)
LIMA (LIM)
LA2457
—
—
—
—
Неизвестно
Играть
19 января 2022
9 Cancun (CUN)
LA2457
LA2457
—
—
—
—
—
Play
18 Январь 2022
5:02
5:02
Приземлился 04:23
18 Jan 20229 Cancun (CUN)
LIMA (LIM)
LA2457
5:02
23:10
23 : 20
04:30
kml CSV Play
9002
18 Jan 2022
Lima (Lim)
9 Cancun (Cun)
LA2456
—
—
—
—
Неизвестно
Играть
9 0013
18 января 2022
4:49 4:49
приземлились 21:35
18 января 2022
Lima (Lim)
Cancun (CUN)
LA2456
4:49
16:35
16:46
22:00
22:00
приземлились 21:35
KML CSV Play
9004
9009
Lima (Lim)
9 Cancun (Cun )
LA2456
—
—
—
—
—
4 Play
3
18 Jan 2022
1:28
30:02
18 января 2022 г.
Икитос (IQT)
Лима (LIM)
LA2231
1:28
9 0991 13:35
13:34
15:20
30:02
9099 kml CSV Play
18 января 2022
1:30
18 Jan 2022
Lima (Lim)
LA2230
LA2230
1:30
10:30
10:35
12:20
kml CSV Play
17 0022
17 Jan 2022
1:16
1:16
17 января 2022
Arequipa (AQP)
Lima (Lim)
LA2124
1:16
20:35
20:32
22:05
Прилетел 21:48
KML CSV Play
2
17 Jan 2022
1:10
1:19
9:19
17 Jan 2022
Lima (Lim)
Arequipa (AQP)
LA2125
1:10
17:55
18:09
19:20
19:20
приземлился 19:19
kml CSV Play
17 Jan 2022
4:35
15:02
17 Jan 2022
Montevideo (MVD)
LIMA (LIM)
LA2421
4:35
10:05
10:27
15:05
Приземлился 15:02
KML CSV Играть
17 января 2022
7 4:10
005 приземлился 08:49
17 января 2022
Lima (Lim)
Montevideo (MVD)
LA2420
4:10
04:35
04:39
09:05
приземлился 08:49
kml CSV Play
17 Jan 2022
1:21
1:21
30:57
17 Jan 2022
Arequipa (AQP)
Lima (Lim)
LA2136
1:21
01:05
01:35
01:35
02:35
поземлился 02:57
KML CSV Play
16 Jan 2022
1:06
Приземлился 00:25
16 января 2022
Лима (LIM)
Арекипа (AQP)
LA2135
LA2135
1:06
23:00
23:19
23:19
00:25
kml CSV Play
16 Jan 2022
0:42
приземлились 22:06
3
16 января 2022
Ayacucho (AYP)
LIMA (LIM)
LA2059
0:42
21:23
21:25
22 : 20
приземлились 22:06
kml CSV Play
16 Jan 2022
0:40
30:07
16 Jan 2022
LIM)
Аякучо (AYP)
LA2058
0:40
19:20
19:27
20:20
приземлился 20:07
kml CSV Play
3
3
16 января 2022
1:33
1:33
30:10
16 Jan 2022
TACNA (TCQ)
Lima (Lim)
LA2164
1:33
16:34
16:37
16:15
18:15
по суше 18:10
KML CSV PLAY
16 Январь 2022
1:29
приземлились 15:44
30:44
16 января 2022
Lima (LIM)
TACNA (TCQ)
LA2165
1:29
14:05
14 :14
15:50
Приземлился 15:44
KML CSV Играть
1 5 января 2022
1:09
приземлился 00:57
9005
15 января 2022
Cusco (CUZ)
9005 LIMA (LIM)
LA2020
1:09
23:50
23:48
01:10
приземлились 00:57
kml CSV Play
15 января 2022
15 1:02
02:35
15 Январь 2022
Lima (Lim)
CUSCO (CUZ)
LA2021
1:02
21:30
21:33
21:45
22:45
приземлился 22:35
KML CSV Play
3
0
15 января 2022
1:10
приземлился 20:02
15 января 2022
CUS Co (Cuz)
Lima (Lim)
LA2062
1:10
18:55
18:52
18:52
20:15
по суше 20:02
KML CSV Play
15 января 2022
1:01
30:49
15 января 2022
Lima (CUZ)
CUSCO (CUZ)
LA2063
1:01
16 : 25
16:48
17:40
17:40
Приземлились 17:49
kml CSV Play
3
15 января 2022
0:39
Приземлились 14:14
15 янв. 2022
Аякучо (AYP)
Лима (LIM)
LA2180
0:39
13:27 1 3
14:20
14:20
Приземлились 14:14
KML CSV PLAY
BFL-1 / A1 Функции, похожих на MCL-1, в качестве селективного антагониста Thbid и Bak
oncogene.Авторская рукопись; Доступно в PMC 2010 Jun 4.
Опубликовано в окончательной редактированной форме AS:
1 Центр для продвинутых Биотехнология и медицина, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси, США
2 Высшая программа по биохимии и молекулярной биологии, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси, США
G Fan
1 Центр передовых биотехнологий и медицины, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси, США
2 Высшая программа по биохимии и молекулярной биологии, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатау ай, Нью-Джерси, США
WX Zong
1 Центр передовых биотехнологий и медицины, UMDNJ—Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси, США
2 Высшая программа по биохимии и молекулярной биологии, UMDNJ—Роберт Медицинская школа Вуда Джонсона, Пискатауэй, Нью-Джерси, США
K Дегенхардт
1 Центр передовых биотехнологий и медицины, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси, США
E White
1 Центр Продвинутая биотехнология и медицина, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси, США
3 Кафедра молекулярной биологии и биохимии, Университет Рутгерса, Нью-Брансуик, Нью-Джерси, США
4 Институт рака Нью-Джерси, Нью-Брансуик, Нью-Джерси, США
C Gélinas
1 Центр передовых биотехнологий и медицины, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси , США
4 Институт рака Нью-Джерси, Нью-Брансуик, Нью-Джерси, США
5 Факультет биохимии, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси, США
1 Центр передовых биотехнологий и медицины, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси, США
2 Высшая программа по биохимии и молекулярной биологии, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси, США
3 Факультет молекулярной Биология и биохимия, Университет Рутгерса, Нью-Брансуик, Нью-Джерси, США
4 Институт рака Нью-Джерси, Нью-Брансуик, Нью-Джерси, США
5 Факультет биохимии, UMDNJ — Медицинская школа Роберта Вуда Джонсона, Пискатауэй, Нью-Джерси , USA
Для переписки: д-р С. Гелинас, CABM и кафедра биохимии, Медицинская школа Роберта Вуда Джонсона, CABM, комната 137, 679 Hoes Lane, Piscataway, NJ 08854-5638, США.ude.sregtur.mbac@sanileg
6 Текущий адрес: Кафедра биологии рака, Медицинская школа Массачусетского университета, Вустер, Массачусетс 01650, США.
7 Текущий адрес: Кафедра молекулярной генетики и микробиологии, SUNY-Stony Brook, Stony Brook, NY 11794-5222, США.
Окончательная отредактированная версия этой статьи доступна на сайте Oncogene См. другие статьи в PMC, в которых цитируется опубликованная статья.
Abstract
Выживающий член семейства Bcl-2 Bfl-1/A1 является транскрипционной мишенью ядерного фактора-κB (NF-κB), который сверхэкспрессируется во многих опухолях человека и является средством, с помощью которого NF-κB ингибирует апоптоз, но его способ действия является спорным.Чтобы лучше понять, как функционирует Bfl-1, мы исследовали его взаимодействие с проапоптотическими мультидоменными белками Bax и Bak, а также с Bh4-белками Bid и tBid. Мы показываем, что в живых клетках Bfl-1 избирательно взаимодействует с Bak и tBid, но не с Bax или Bid. Взаимодействие Bfl-1/Bak является функциональным, поскольку Bfl-1 подавляет индуцированный стауроспорином (STS) апоптоз в клетках дикого типа и Вах-дефицитных клетках, но не в клетках Bak-/-. Мы также показываем, что Bfl-1 косвенно блокирует активацию Bax, индуцированную фактором некроза опухоли-α (TNFα), через связь с tBid.Делеция С-конца снижала взаимодействие Bfl-1 с Bak и tBid и снижала его способность подавлять Bak- и tBid-опосредованную гибель клеток. Эти данные показывают, что Bfl-1 использует различные механизмы для подавления апоптоза в зависимости от стимула. Bfl-1 связывается с tBid, чтобы предотвратить активацию проапоптотических Bax и Bak, а также напрямую взаимодействует с Bak, чтобы противодействовать опосредованной Bak гибели клеток, подобно Mcl-1. Таким образом, часть защитной функции NF-κB заключается в индукции Mcl-1-подобной активности путем усиления Bfl-1.
Взаимодействия между белками семейства Bcl-2 контролируют клеточную судьбу в ответ на сигналы, индуцирующие гибель ). Белки, содержащие только проапоптотический Bh4 (Bid, Bad, Bim, Puma, Noxa, Hrk, Bmf, Nbk/Bik), инициируют передачу сигналов апоптоза в ответ на вызывающие смерть стимулы. Многодоменные белки Bax и Bak являются важными эффекторами апоптоза, и клетки, лишенные обоих белков, невосприимчивы к апоптозу, индуцированному широким спектром стимулов.Напротив, антиапоптотические члены семейства Bcl-2 Bcl-2, Bcl-x L , Bfl-1/A1, Mcl-1 и Bcl-w защищают клетки от апоптоза. Некоторые белки, содержащие только Bh4, такие как tBid и Bim, запускают конформационную активацию Bax и Bak, в то время как другие, такие как Bad, Noxa и Bik, секвестрируют антиапоптотические члены семейства Bcl-2 и нейтрализуют их защитную активность (Letai et al. , 2002). ; Chen и др., 2005; Kuwana и др. , 2005).
Апоптоз может быть инициирован внешним путем, включающим активацию рецепторов смерти, или внутренним путем, запускаемым различными формами клеточного стресса.Во внутриклеточном пути апоптотические сигналы сходятся на митохондриях, чтобы вызвать высвобождение цитохрома C (cyto C ) в цитозоль, вызывая активацию каспазы и гибель клетки. Во внешнем пути лиганды, такие как фактор некроза опухоли-α (TNFα), вызывают опосредованную рецептором смерти активацию каспазы-8, которая расщепляет Bid до его активной формы tBid. В свою очередь, tBid индуцирует конформационную активацию и олигомеризацию Bax и Bak, высвобождение митохондриального цито C , активацию каспазы и апоптоз (Perez and White, 2000; Wei et al., 2000). В то время как антиапоптотические Bcl-2 и Bcl-x L могут связываться как с Bid, так и с tBid, Mcl-1 ингибирует апоптоз, индуцированный рецептором смерти, путем селективного связывания с tBid, но не с Bid (Cheng et al. , 2001; Clohessy). и др. , 2006). Более того, Mcl-1 изолирует эндогенный Bak, но не Bax в здоровых клетках, и блокирует Bak-опосредованную гибель клеток, а Bcl-2 этого не делает (Cuconati et al. , 2003; Willis et al. , 2005). Аденовирусный гомолог Bcl-2 E1B 19K препятствует гибели клеток, связываясь непосредственно с Bax и Bak, но не с tBid или Bid (Oltvai et al., 1993; Перес и Уайт, 2000 г.; Куконати и др. , 2002; Cuconati and White, 2002), но как другие антиапоптотические члены семейства Bcl-2, такие как Bfl-1, блокируют апоптоз, не совсем ясно.
Bfl-1/A1 является транскрипционной мишенью ядерного фактора-κB (NF-κB), который подавляет апоптоз в ответ на несколько вызывающих смерть стимулов, включая TNFα и стауроспорин (STS; Karsan et al. , 1996; D ‘Sa-Eipper and Chinnadurai, 1998; Grumont и др. , 1999; Lee и др., 1999; Ван и др. , 1999; Зонг и др. , 1999; Шомоджи и др. , 2001). Bfl-1/A1 не имеет значительной гомологии с доменом Bh5, который необходим для защитной активности Bcl-2 и Bcl-x L , а его С-концевая область содержит заряженные остатки и вряд ли будет действовать как трансмембранный домен, подобный которые прикрепляют Bcl-2 и Bcl-x L к митохондриальным мембранам (Nguyen et al. , 1993). Это предполагает, что Bfl-1 может действовать иначе, чем Bcl-2 и Bcl-x L , блокируя апоптоз, и имеются противоречивые сообщения относительно его способности связываться с Bax, Bak, Bid и tBid. Сообщалось, что человеческий Bfl-1 взаимодействует с Bid в временно трансфицированных клетках и с рекомбинантным tBid in vitro , но не с Bax или Bak (Werner et al. , 2002). Было описано, что Bfl-1/A1 связывается с Bax в двухгибридных дрожжах или путем совместной иммунопреципитации in vitro транслированных белков (Sedlak et al. , 1995; Zhang et al. , 2000). Другие исследователи не обнаружили связи между A1 мыши и Bax в котрансфицированных клетках, но A1 взаимодействовал с Bak в этих условиях (Holmgreen et al., 1999). Здесь мы исследовали связь Bfl-1 с эндогенными Bax, Bak, Bid и tBid в живых клетках и изучили, как эти взаимодействия связаны с его защитной активностью в ответ на внешние и внутренние стимулы, вызывающие смерть.
Результаты
Поскольку Bfl-1 гиперэкспрессируется при многих видах рака, мы экспрессировали Bfl-1 в клетках MCF-7 и подтвердили его способность подавлять высвобождение митохондриального цито C при активации TNFα внешнего сигнального каскада смерти. Bfl-1 был помечен с N-конца зеленым флуоресцентным белком (GFP), поскольку коммерческие антитела не смогли успешно распознать Bfl-1 человека. GFP-Bfl-1 локализуется в митохондриях и околоядерной области, перекрывая эндогенный цито C (). В то время как TNFα провоцировал высвобождение цито C в экспрессирующих GFP клетках, цито C оставался митохондриально локализованным в GFP-Bfl-1-положительных клетках, что согласуется с его защитной активностью в отношении TNFα (Karsan et al. , 1996; Wang ). и другие., 1999; Зонг и др. , 1999). Мутант GFP-Bfl-1ΔC, лишенный С-концевых 24 аминокислот, распространялся по всей клетке, включая ядро, подобно GFP, но все еще подавлял TNFα-индуцированное высвобождение цито C . Высвобождение Cyto C и апоптоз критически зависят от конформационной активации и олигомеризации Bax и Bak, которые ингибируются антиапоптотическими Bcl-2 и Bcl-x L . Точно так же GFP-Bfl-1 и Bfl-1ΔC подавляли TNFα-индуцированную конформационную активацию Bax, как видно из антитела, специфичного к N-концу Bax, которое маскируется, когда Bax неактивен, и экспонируется при активации Bax (; Desagher et al. ., 1999; Перес и Уайт, 2000). Напротив, Bax активировался в окружающих клетках, в которых отсутствовала значительная экспрессия GFP-Bfl-1. Таким образом, Bfl-1 и Bfl-1ΔC подавляют активацию Bax в ответ на TNFα.
Bfl-1 и Bfl-1ΔC подавляют TNα-индуцированное высвобождение цито C . Иммунофлуоресценция клеток MCF-7, трансфицированных GFP ( a – f ), GFP-Bfl-1 ( g – n ) или –Bfl1ΔC ( o–v ) с ( d 9 F , K — N ; с — S — S — S — S ) или без ( C — г , г — j , o — R ) Лечение TKFα для 8 h, с использованием анти-цито C антитела ( c , f , i , m , q , u ). Клетки также анализировали на флуоресценцию GFP ( b , e , h , l , p , t ). Цито С , цитохром С ; GFP, зеленый флуоресцентный белок; TNFα, фактор некроза опухоли-α.
Bfl-1 и BfI-1ΔC подавляют TNFα-индуцированную конформационную активацию Bax. Иммунофлуоресценция активированного Bax в клетках MCF-7, трансфицированных GFP ( a – d ), GFP-Bfl-1 ( e – h ) или -Bfl-1ΔC ( i – l ) С ( C и D , г и ч 9175 и ч , к и к и л и л ) или без ( A и B , E и F , I и j ) Обработка TNFα с использованием антитела против BaxNT ( b , d , f , h , j , l ). Клетки также анализировали на флуоресценцию GFP (—, -с-, —, —, -1-, —). Звездочками отмечены GFP-Bfl-1 или -Bfl-1 AC-положительные клетки, которые не показали активации Bax.
Члены семейства антиапоптотических Bcl-2 блокируют апоптоз, взаимодействуя с проапоптотическими белками, состоящими только из Bh4, и/или с мультидоменными белками Bax и Bak. Bcl-2 и E1B 19K взаимодействуют с Bax, блокируя его олигомеризацию и апоптоз (Oltvai et al. , 1993; Perez and White, 2000; Cuconati and White, 2002; Cuconati et al., 2002), но связь Bfl-1 с Bax была предметом противоречивых сообщений (Sedlak et al. , 1995; Holmgreen et al. , 1999; Zhang et al. , 2000; Werner ). и др. , 2002). Мы спросили, противодействует ли Bfl-1 активации Bax посредством прямого взаимодействия с экстрактами 3-((3-холамидопропил)диметиламмонио)-1-пропансульфоната (CHAPS), поскольку другие детергенты могут влиять на конформацию Bax и его взаимодействие с Bcl-2 (Hsu and Youle). , 1998). Хотя Bfl-1 и Bfl-1ΔC ко-иммунопреципитируются со сверхэкспрессированным FLAG-Bax (1), ни один из них не связан ни с эндогенным Bax, активированным TNFα/циклогексимидом (CHX), ни с неактивным Bax в клетках, обработанных CHX, несмотря на эффективную иммунопреципитацию Bax (2).Обратная иммунопреципитация эндогенного Bax с помощью GFP-Bfl-1 или 2×Myc-Bfl-1 также не удалась, в отличие от контрольной иммунопреципитации эндогенного Bax с помощью Bak (14). Т.о., хотя Bfl-1 может ассоциироваться со сверхэкспрессией Bax, комплексов с эндогенным Bax не наблюдалось, что свидетельствует о том, что он противодействует активации Bax, взаимодействуя с факторами, действующими выше каскада TNF α.
Bfl-1 и Bfl-1ΔC могут взаимодействовать со сверхэкспрессированным Bax, но не могут связываться с эндогенным Bax независимо от статуса активации Bax.( a ) Совместная иммунопреципитация GFP-Bfl-1 или -Bfl-1ΔC со сверхэкспрессией FLAG-Bax в экстрактах CHAPS из котрансфицированных клеток HeLa. ( b ) Иммунопреципитация 2×Myc-Bfl-1 или -Bfl-1ΔC с эндогенным Bax в клетках HeLa, обработанных TNFα плюс CHX или только CHX, с использованием антител против Bax-loop и иммуноблоттинга с анти-Myc или -BaxN20 антитело. Lys: 1/10 вводного лизата. CHX, циклогексимид; GFP, зеленый флуоресцентный белок; TNFα, фактор некроза опухоли-α.
Bfl-1 избирательно взаимодействует с эндогенным Bak, но не с Bax, и подавляет апоптоз в клетках iBMK дикого типа и Bax-дефицитных, но не в Bak-дефицитных iBMK.( a ) Коиммунопреципитация эндогенного Bax или Bak с GFP-Bfl-1, эндогенным Bak или 2×Myc-Bfl-1 в клетках HeLa с обработкой TNFα/CHX или без нее для активации Bax и Bak. NT: нетрансфицированный. Линия разграничивает разные экспозиции одного и того же пятна. ( b ) Коиммунопреципитацию эндогенного Bax или Bak с 2×Myc-Bfl-1 по сравнению с FLAG-Mcl-1 проводили, как в панели ( a ). ( c ) Иммунопреципитация эндогенного Bak с помощью 2×Myc-Bfl-1, -Bfl-1ΔC или FLAG-Mcl-1. ( d ) Иммортализованные iBMK дикого типа, Bak-/-, Bax-/- и Bax-/-Bak-/- с двойным нокаутом, трансфицированные EGFP (заштрихованы), 2×Myc-Bfl-1 (черные), 2×Myc-Bfl-1ΔC (белый) или FLAG-Mcl-1 (точечный) обрабатывали STS в течение 24 ч или оставляли без обработки. Среднюю выживаемость в трех экспериментах определяли по исключению трипанового синего и нормализовали по эффективности трансфекции. CHX, циклогексимид; GFP, зеленый флуоресцентный белок; iBMK, бессмертная почка мышиного детеныша; STS, стауроспорин; TNFα, фактор некроза опухоли-α.
В противоположность этому GFP-Bfl-1 и 2×Myc-Bfl-1 сильно взаимодействовали с эндогенным Bak в необработанных клетках, подобно Mcl-1, а также с TNFα-активированным Bak (). Пониженные уровни Bak, иммунопреципитированного с 2×Myc-Bfl-1 в клетках, обработанных TNFα/CHX, согласуются с более низкими уровнями 2×Myc-Bfl-1 в этих условиях, что соответствует его обороту (Kucharczak et al. , 2005). ; Герольд и др. , 2006). Взаимодействие Bfl-1ΔC с Bak было значительно слабее, чем у Bfl-1 ().Таким образом, Bfl-1 ассоциирует с неактивной и активной формами эндогенного Bak, тогда как сниженное взаимодействие Bfl-1ΔC, скорее всего, является результатом его делокализации из митохондрий.
Избирательное взаимодействие Bfl-1 с эндогенным Bak является функциональным, поскольку он подавляет внутренний STS-индуцированный апоптоз в клетках эпителия почек бессмертных мышей дикого типа и Bax-дефицитных (iBMK), но не в Bak-дефицитных iBMK, которые экспрессируют Bax, но не Бак (; Degenhardt et al. , 2002). Два независимых клона iBMK каждого генотипа дали сходные результаты (не показаны).Поведение Bfl-1 было поразительно похоже на Mcl-1, который также взаимодействует с Bak, но не с Bax (Cuconati et al. , 2003; Gelinas and White, 2005; Willis et al. , 2005). Как и ожидалось, iBMK с двойным нокаутом, лишенные Bax и Bak, были очень устойчивы к апоптозу. Bfl-1ΔC был менее защитным, чем Bfl-1, в отношении Bax-/- и iBMK дикого типа, что согласуется с его сниженной связью с Bak. Как и в случае с Bfl-1, Bfl-1ΔC не способен подавлять апоптоз в клетках Bak-/-. Это показало, что селективное взаимодействие Bfl-1 с Bak, но не с Bax, коррелирует с подавлением Bak-, но не Bax-опосредованного апоптоза, подобно Mcl-1.
TNFα-индуцированный процессинг Bid в tBid приводит к активации Bax и Bak. Поскольку одновременная инактивация Bax и Bak необходима для противодействия смерти, вызванной TNFα (Degenhardt et al. , 2002), а Bfl-1 не взаимодействует с эндогенным Bax, мы спросили, связана ли его способность подавлять активацию Bax с взаимодействием с Bid и /или ставка. Сверхэкспрессия Bid-Myc или tBid-Myc коиммунопреципитировала GFP-Bfl-1 и Bfl-1ΔC (), как наблюдалось ранее (Werner et al. , 2002).Однако оба не смогли ассоциироваться с эндогенным Bid, несмотря на эффективную иммунопреципитацию 2×Myc-Bfl-1 и Bfl-1ΔC (10). Меньшее количество Bfl-1 по сравнению с Bfl-1ΔC во входных дорожках согласуется с конститутивным оборотом Bfl-1, который включает его С-конец (Kucharczak et al., 2005; Herold et al. , 2006). Результаты заметно отличались, когда мы исследовали ассоциацию Bfl-1 с эндогенным tBid в клетках, обработанных TNFα/CHX, чтобы способствовать процессингу Bid в tBid (). Хотя эндогенный tBid трудно обнаружить из-за его короткого периода полувыведения (Breitschopf et al., 2000), было ясно видно взаимодействие с 2×Myc-Bfl-1 или Bfl-1ΔC, особенно при более длительном воздействии. В поддержку функционального взаимодействия Bfl-1 и Bfl-1ΔC подавляли апоптоз, индуцированный tBid, хотя Bfl-1ΔC был несколько менее эффективен, чем Bfl-1 (12). Это показывает, что подобно его дифференциальному взаимодействию с эндогенным Bak, но не с Bax, Bfl-1 избирательно связывается с эндогенным tBid, но не с Bid, и предполагает, что это взаимодействие важно для подавления внешнего апоптотического каскада, инициируемого TNFα.
Bfl-1 и Bfl-1ΔC избирательно взаимодействуют с эндогенным tBid, но не с Bid в клетках, обработанных TNFα, и подавляют апоптоз, индуцированный tBid. ( a ) Иммунопреципитация/вестерн-блоттинг GFP-Bfl-1 или -Bfl-1ΔC со сверхэкспрессией Bid-Myc или tBid-Myc в клетках HeLa. ( b ) Коиммунопреципитация эндогенного Bid или tBid с 2×Myc-Bfl-1 или -Bfl-1ΔC в клетках HeLa, обработанных только TNFα/CHX или CHX. Показана более длительная экспозиция. Звездочками отмечены позиции эндогенных Bid и tBid; над Bid в лизатах появлялась неспецифическая полоса.( c ) GFP-Bfl-1 или -Bfl-1ΔC подавляют вызванную tBid гибель клеток в клетках MCF-7, котрансфицированных tBid-FLAG или pcDNA3 вместе с CMV-β-gal. Показано среднее из трех экспериментов. ( d ) Модель, иллюстрирующая избирательное взаимодействие Bfl-1 с tBid и Bak для подавления TNFα- и Bak-зависимого апоптоза, а также сходство его механизма действия с Mcl-1. CHX, циклогексимид; GFP, зеленый флуоресцентный белок; TNFα, фактор некроза опухоли-α.
Обсуждение
Расхождение последовательностей между Bfl-1 и другими антиапоптотическими членами семейства Bcl-2 вызвало вопросы относительно механизма его действия. Мы демонстрируем, что в функциональном контексте живых клеток, где Bfl-1 ингибирует апоптоз, Bfl-1 избирательно связывается с эндогенными tBid и Bak, но не с Bid или Bax. В то время как Bfl-1 противодействовал активации Bax в ответ на внешнюю передачу сигнала о смерти с помощью TNFα, он не смог подавить Bax-опосредованную гибель клеток в ответ на внутреннюю передачу сигнала о смерти, которая не зависит от tBid в клетках Bak-/-, но эффективно блокировал STS-индуцированный апоптоз. в клетках Bax-/-, которые экспрессируют эндогенный Bak. Это указывает на то, что конститутивное взаимодействие Bfl-1 с Bak важно для подавления Bak-опосредованного апоптоза, тогда как его связь с tBid, скорее всего, ответственна за блокирование активации Bax в ответ на стимуляцию рецептора смерти.
Преобразование Bid в tBid усиливает сигнализацию рецептора смерти и приводит к активации Bax и Bak. В некоторых исследованиях предполагается, что tBid действует непосредственно на митохондрии, способствуя высвобождению цито C в отсутствие Bax и/или Bak (Epand et al., 2002; Grinberg et al. , 2002). Однако исследования нокаута показали, что tBid, скорее всего, действует путем активации Bax и Bak, поскольку клетки, лишенные обоих факторов, устойчивы к апоптозу (Wei et al. , 2001; Degenhardt et al., 2002). Наши данные подтверждают идею о том, что ассоциация Bfl-1 с tBid, скорее всего, ответственна за подавление активации Bax, поскольку Bfl-1 не смог связать эндогенный Bax в клетках, обработанных TNF. Это согласуется с сообщениями, показывающими, что, хотя Bcl-2 и Bcl-x L могут напрямую ассоциироваться с Bax, их взаимодействие с tBid наиболее важно для их защитной функции в каскаде TNF (Cheng et al. , 2001; Werner ). и др. , 2002; Yi и др. , 2003).
Важно отметить, что подобно Mcl-1, Bfl-1 взаимодействует с Bak-опосредованным апоптозом и подавляет его, и это взаимодействие не зависит от активации Bak (Cuconati et al. , 2003; Willis et al. , 2005). Недавняя работа предполагает, что они также демонстрируют поразительно сходное сродство к пептидам Bh4 из некоторых факторов, содержащих только Bh4, которое отличается от такового у Bcl-2, Bcl-x L и Bcl-w (Chen et al., 2005), хотя некоторые обнаружили иное (Certo et al. , 2006).Наш вывод о том, что Bfl-1 физически и функционально взаимодействует с Bak в живых клетках, отличается от других, которым не удалось увидеть взаимодействие между , транслируемым in vitro Bfl-1, и Bak в присутствии митохондрий печени мыши (Werner et al. , 2002). ). Поскольку рекомбинантный tBid был включен в эти анализы, а ассоциация tBid с Mcl-1 может вытеснять Bak из комплексов Mcl-1-Bak (Clohessy et al. , 2006), tBid также может диссоциировать Bfl-1 от Bak, тем самым препятствуя обнаружению комплексов Bfl-1-Bak в этих анализах.
Поскольку гидрофобный С-конец мультидоменных белков семейства Bcl-2 участвует в локализации мембраны, а заряженные остатки присутствуют на С-конце Bfl-1, Bfl-1 может быть белком периферической мембраны, который слабо связан с митохондриями и может зависеть от на взаимодействие с митохондриальными белками для локализации в этой органелле. Хотя мы не исключаем возможности того, что Bfl-1ΔC сохраняет некоторую локализацию в митохондриях, его измененная локализация согласуется с его сниженной ассоциацией с эндогенным Bak, которая постоянно обнаруживается в митохондриях, и его сниженной способностью эффективно подавлять Bak-зависимый апоптоз. Это может объяснить, почему в недавней работе с использованием Bfl-1ΔC-подобной формы A1, лишенной 20 C-концевых аминокислот, не удалось обнаружить взаимодействие с Bak (Chen et al. , 2005; Willis et al. , 2005). ). Ослабленная ассоциация Bfl-1ΔC с Bak может также прояснить, почему делеция C-конца Bfl-1 снижает его защитную активность в отношении p53-индуцированного апоптоза, в котором Bak играет важную роль (D’Sa-Eipper and Chinnadurai, 1998). В совокупности эти находки показывают, что эффективное взаимодействие Bfl-1/Bak может быть важным средством для Bfl-1 противодействовать внутренней передаче сигналов смерти, которая зависит от Bak и не передает сигналы через tBid.
В целом наши данные показывают, что механизм действия Bfl-1 подобен действию Mcl-1, который изолирует Bak в неактивных комплексах и взаимодействует с tBid для подавления апоптоза, опосредованного рецептором смерти (; Oltvai et al. , 1993; Perez and White, 2000; Cuconati and White, 2002; Cuconati и др., 2002, 2003; Clohessy и др. , 2006). Хотя Bfl-1 и Mcl-1 используют сходные средства для подавления апоптоза, они, вероятно, действуют неповторяющимся образом в разных клетках и в ответ на разные стимулы, поскольку они находятся под разным транскрипционным контролем, а Mcl-1 не является Мишень NF-κB (Michels et al., 2005). Bfl-1 сверхэкспрессируется во многих опухолях человека, в том числе в некоторых, в которых конститутивная активация NF-κB необходима для выживания и химиорезистентности. Таким образом, функциональное взаимодействие Bfl-1 с tBid и Bak может способствовать его роли в онкогенезе и резистентности к терапии. В совокупности эти исследования позволяют по-новому взглянуть на механизмы, с помощью которых Bfl-1 противодействует внешним и внутренним каскадам сигналов смерти, и позволяют предположить, что подходы к специфической блокировке взаимодействия Bfl-1 с tBid и/или Bak могут улучшить реакцию клеток, экспрессирующих Bfl-1. опухолевых клеток к противораковому лечению. Это может быть важно, поскольку ингибирование Mcl-1 необходимо для сенсибилизации некоторых опухолей к низкомолекулярным миметикам Bh4 ABT-737, которые противодействуют Bcl-x L , Bcl-2 и Bcl-w (Konopleva et al. ). , 2006; Letai, 2006; van Delft и др. , 2006). Если Bfl-1 представляет собой NF-κB-индуцированный заменитель Mcl-1, антагонистическая функция Bfl-1 может быть необходима для терапии рака и, возможно, является одним из средств, с помощью которых ингибиторы NF-κB провоцируют апоптоз раковых клеток.
Материалы и методы.
Иммунофлуоресценция.
Иммунопреципитация
Клетки HeLa, котрансфицированные FLAG-Bax, Bid-Myc или tBid-Myc, были дополнены ингибитором каспаз z-Val-Ala-Asp(OMe)-FMK (zVAD-fmk). Взаимодействие с эндогенными факторами анализировали в экстрактах CHAPS из клеток HeLa, обработанных TNFα (10 нг мл -1 ) плюс CHX (30 мкг мл -1 ) или только CHX в течение 6 ч с использованием анти-FLAG, -Myc, -Bak или -Bax-loop антитела и иммуноблотинг для Myc, GFP, BaxNT, BakNT, BaxN20 или Bid.
Анализы апоптоза
Иммортализованные клеточные линии BMK, трансфицированные усиленным зеленым флуоресцентным белком (EGFP), Bfl-1, Bfl-1ΔC или Mcl-1, не обрабатывали или обрабатывали STS (1.5 мкМ) в течение 24 часов. Выживаемость определяли по исключению трипанового синего и нормализовали по эффективности трансфекции. Определяли выживаемость β-галактозидазо-положительных клеток MCF-7, котрансфицированных GFP-Bfl-1 или -Bfl-1ΔC и tBid-FLAG или pcDNA3 вместе с pCMV-β-gal.
Дополнительные детали эксперимента доступны в дополнительной информации.
Благодарности
Мы благодарим R Sundararajan, D Perez и N Gupta за обсуждения. Эта работа была поддержана грантом NIH CA083937, Фондом Шарлотты Гейер и Фондом UMDNJ.MJS был частично поддержан грантом GM08360 для подготовки докторантов NIH.
Ссылки
Breitshopf K, Zeiher AM, Dimmeler S. Опосредованная убиквитином деградация проапоптотически активной формы bid. Функциональное последствие индукции апоптоза. Дж. Биол. Хим. 2000; 275:21648–21652. [PubMed] [Google Scholar]
Certo M, Del Gaizo Moore V, Nishino M, Wei G, Korsmeyer S, Armstrong SA, et al. Митохондрии, инициированные сигналами смерти, определяют клеточную зависимость от антиапоптотических членов семейства BCL-2.Раковая клетка. 2006; 9: 351–365. [PubMed] [Google Scholar]
Chen L, Willis SN, Wei A, Smith BJ, Fletcher JI, Hinds MG, et al. Дифференциальное нацеливание на белки Bcl-2, способствующие выживанию, с помощью их лигандов, состоящих только из Bh4, делает возможной комплементарную апоптотическую функцию. Мол Ячейка. 2005; 17: 393–403. [PubMed] [Google Scholar]
Cheng EH, Wei M, Weiler S, Flavell RA, Mak TW, Lindsten T, et al. BCL-2, BCL-XL изолируют молекулы, содержащие только домен Bh4, предотвращая BAX- и BAK-опосредованный митохондриальный апоптоз. Мол Ячейка.2001; 8: 705–711. [PubMed] [Google Scholar]
Clohessy JG, Zhuang J, de Boer J, Gil-Gomez G, Brady HJ. Mcl-1 взаимодействует с укороченным Bid и ингибирует его индукцию высвобождения цитохрома c и его роль в рецептор-опосредованном апоптозе. Дж. Биол. Хим. 2006; 281:5750–5759. [PubMed] [Google Scholar]
Cory S, Huang DC, Adams JM. Семейство Bcl-2: роль в выживании клеток и онкогенезе. Онкоген. 2003; 22:8590–8607. [PubMed] [Google Scholar]
Куконати А., Дегенхардт К., Сундарараджан Р., Аншель А., Уайт Э.Bak и Bax ограничивают репликацию аденовирусов посредством индукции апоптоза. Дж Вирол. 2002; 76: 4547–4558. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Cuconati A, Mukherjee C, Perez D, White E. Реакция на повреждение ДНК и разрушение MCL-1 инициируют апоптоз в инфицированных аденовирусом клетках. Гены Дев. 2003; 17: 2922–2932. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Cuconati A, White E. Вирусные гомологи Bcl-2: роль апоптоза в регуляции вирусной инфекции. Гены Дев.2002; 16: 2465–2478. [PubMed] [Google Scholar]
D’Sa-Eipper C, Chinnadurai G. Функциональная диссекция Bfl-1, гомолога Bcl-2: антиапоптозная активность, сотрудничество с онкогенами и пролиферация клеток. Онкоген. 1998;16:3105–3114. [PubMed] [Google Scholar]
Дегенхардт К., Сундарараджан Р., Линдстен Т., Томпсон С., Уайт Э. Бакс и Бак независимо способствуют высвобождению цитохрома С из митохондрий. Дж. Биол. Хим. 2002; 277:14127–14134. [PubMed] [Google Scholar]
Desagher S, Osen-Sand A, Nichols A, Eskes R, Montessuit S, Lauper S, et al.Индуцированное Bid конформационное изменение Bax отвечает за высвобождение митохондриального цитохрома c во время апоптоза. Джей Селл Биол. 1999; 144: 891–901. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Gélinas C, White E. Белки, содержащие только Bh4, в контроле: специфичность регулирует MCL-1 и BAK-опосредованный апоптоз.Гены Дев. 2005;19:1263–1268. [PubMed] [Google Scholar]
Гринберг М. , Сариг Р., Зальцман Ю., Фрумкин Д., Грамматикакис Н., Реувени Э. и др. tBID гомоолигомеризуется в митохондриальной мембране, вызывая апоптоз. Дж. Биол. Хим. 2002; 277:12237–12245. [PubMed] [Google Scholar]
Grumont RJ, Rourke IJ, Gerondakis S. Rel-зависимая индукция транскрипции A1 необходима для защиты B-клеток от апоптоза, вызванного лигированием антигенных рецепторов. Гены Дев. 1999; 13:400–411. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Herold MJ, Zeitz J, Pelzer C, Kraus C, Peters A, Wohlleben G, et al.Стабильность и антиапоптотическая функция А1 контролируются его С-концом. Дж. Биол. Хим. 2006; 281:13663–13671. [PubMed] [Google Scholar]
Holmgreen SP, Huang DC, Adams JM, Cory S. Выживаемость гомологов Bcl-2 Bcl-w и A1 лишь частично коррелирует с их способностью связывать проапоптотических членов семейства. Смерть клеток 1999; 6: 525–532. [PubMed] [Google Scholar]
Хсу Ю.Т., Юле Р.Дж. Bax в тимусе мышей представляет собой растворимый мономерный белок, который демонстрирует различные конформации, индуцированные детергентом. Дж. Биол. Хим. 1998; 273:10777–10783. [PubMed] [Google Scholar]
Karsan A, Yee E, Harlan JM, et al. Гибель эндотелиальных клеток, индуцированная фактором некроза опухоли-альфа, ингибируется членом семейства Bcl-2, A1. Дж. Биол. Хим. 1996; 271:27201–27204. [PubMed] [Google Scholar]
Коноплева М., Контрактор Р., Цао Т., Самудио И., Руволо П.П., Китада С. и др. Механизмы чувствительности и устойчивости к апоптозу к миметику Bh4 ABT-737 при остром миелоидном лейкозе. Раковая клетка. 2006; 10: 375–388.[PubMed] [Google Scholar]
Kucharczak JF, Simmons MJ, Duckett CS, Gelinas C. Опосредованный конститутивными протеасомами оборот Bfl-1/A1 и его процессинг в ответ на активацию рецептора TNF в клетках FL5.2 pro-B convert это в фактор предсмерти. Смерть клеток 2005; 12:1225–1239. [PubMed] [Google Scholar]
Kuwana T, Bouchier-Hayes L, Chipuk JE, Bonzon C, Sullivan BA, Green DR, et al. Домены Bh4 либо сенсибилизируют, либо активируют митохондриальный апоптоз, выступая в качестве прототипа противораковой терапии. Мол Ячейка. 2005; 17: 525–535. [PubMed] [Google Scholar]
Lee HH, Dadgostar H, Cheng Q, Shu J, Cheng G. Опосредованная NF-kappaB активация Bcl-x и Bfl-1/A1 необходима для передачи сигналов выживания CD40 в B-лимфоцитах. Proc Natl Acad Sci USA. 1999;96:9136–9141. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Летай А. Восстановление смертного приговора раку. Раковая клетка. 2006; 10: 343–345. [PubMed] [Google Scholar]
Letai A, Bassik MC, Walensky LD, Sorcinelli MD, Weiler S, Korsmeyer SJ.Отдельные домены Bh4 либо сенсибилизируют, либо активируют митохондриальный апоптоз, выступая в качестве прототипа противораковой терапии. Раковая клетка. 2002; 2: 183–192. [PubMed] [Google Scholar]
Michels J, Johnson PW, Packham G. Mcl-1. Int J Biochem Cell Biol. 2005; 37: 267–271. [PubMed] [Google Scholar]
Нгуен М., Миллар Д.Г., Йонг В.В., Корсмейер С.Дж., Шор Г.К. Нацеливание Bcl-2 на наружную мембрану митохондрий с помощью COOH-концевой сигнальной якорной последовательности. Дж. Биол. Хим. 1993; 268:25265–25268. [PubMed] [Google Scholar]
Олтвай З.Н., Миллиман С.Л., Корсмейер С.Дж.Bcl-2 гетеродимеризует in vivo с консервативным гомологом Bax, который ускоряет запрограммированную гибель клеток. Клетка. 1993; 74: 609–619. [PubMed] [Google Scholar]
Перес Д., Уайт Э. ФНО-альфа сигнализирует об апоптозе через конформационное изменение Bax, зависящее от бид, которое ингибируется E1B 19K. Мол Ячейка. 2000; 6: 53–63. [PubMed] [Google Scholar]
Sedlak TW, Oltvai ZN, Yang E, Wang K, Boise LH, Thompson CB, et al. Несколько членов семейства Bcl-2 демонстрируют селективную димеризацию с Bax.Proc Natl Acad Sci USA. 1995; 92: 7834–7838. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Somogyi RD, Wu Y, Orlofsky A, Prystowsky MB. Временная экспрессия члена семейства Bcl-2, A1-a, приводит к ядерной локализации и устойчивости к стауроспорин-индуцированному апоптозу. Смерть клеток 2001; 8: 785–793. [PubMed] [Google Scholar]
van Delft MF, Wei AH, Mason KD, Vandenberg CJ, Chen L, Czabotar PE, et al. Миметик Bh4 ABT-737 нацеливается на селективные белки Bcl-2 и эффективно индуцирует апоптоз через Bak/Bax, если Mcl-1 нейтрализуется.Раковая клетка. 2006; 10: 389–399. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Wang CY, Guttridge DC, Mayo MW, Baldwin AS., Jr NF-kappaB индуцирует экспрессию гомолога Bcl-2 A1/Bfl-1 для преимущественного подавления индуцированного химиотерапией апоптоз. Мол Селл Биол. 1999;19:5923–5929. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Wei MC, Lindsten T, Mootha VK, Weiler S, Gross A, Ashiya M, et al. tBID, мембранно-направленный лиганд смерти, олигомеризует BAK с высвобождением цитохрома c .Гены Дев. 2000;14:2060–2071. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Wei MC, Zong WX, Cheng EH, Lindsten T, Panoutsakopoulou V, Ross AJ, et al. Проапоптотические BAX и BAK: необходимые ворота к митохондриальной дисфункции и гибели. Наука. 2001; 292: 727–730. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Werner AB, de Vries E, Tait SWG, Bontjer I, Borst J. Член семьи Bcl-2 секвестрирует Bfl-1/A1 укорочено, чтобы предотвратить его сотрудничество с проапоптотическим Bak или Бакс.Дж. Биол. Хим. 2002; 277: 22781–22788. [PubMed] [Google Scholar]
Willis SN, Chen L, Dewson G, Wei A, Naik E, Fletcher JI, et al. Проапоптотический Bak секвестрируется Mcl-1 и Bcl-xL, но не Bcl-2, до тех пор, пока не будет вытеснен белками, состоящими только из Bh4. Гены Дев. 2005;19:1294–1305. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Yi X, Yin XM, Dong Z. Ингибирование Bid-индуцированного апоптоза с помощью Bcl-2. Инсерция tBid, транслокация Bax и олигомеризация Bax/Bak подавлены. Дж. Биол. Хим. 2003; 278:16992–16999.[PubMed] [Google Scholar]
Zhang H, Cowan-Jacob SW, Simonen M, Greenhalf W, Heim J, Meyhack B. Структурная основа Bfl-1 для его взаимодействия с Bax и антиапоптотического действия в клетках млекопитающих и дрожжей . Дж. Биол. Хим. 2000; 275:11092–11099. [PubMed] [Google Scholar]
Zong WX, Edelstein LC, Chen C, Bash J, Gélinas C. Гомолог Bcl-2 для выживания Bfl-1/A1 является прямой мишенью транскрипции NF-κB, который блокирует апоптоз, индуцированный TNFα. . Гены Дев. 1999; 13: 382–387. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Можете ли вы оспорить вмешательство в передачу в НФЛ? Правила повторов и многое другое об OPI, DPI в 2020 году
Возможно, есть люди, которые тоскуют по дням, когда помехи при пасах в атаке и защите были фолами, подпадающими под действие системы просмотра повторов НФЛ.Некоторым, как говорится, просто нравится смотреть, как горит мир.
По ходу сезона НФЛ 2020 года не трудитесь кричать в телевизор, чтобы тренер вашей любимой команды бросил свой флаг вызова или чтобы официальные лица игры направились к монитору мгновенного повтора, когда вы видите то, что, по вашему мнению, пропущено. передать вызов помех. Эти штрафы больше не подлежат пересмотру в НФЛ.
ДЕКУРСИ: Как НФЛ потерпела неудачу в обзоре повтора PI
Сага о просмотре повтора помех НФЛ была катастрофическим, годичным экспериментом, родившимся из печально известного пропущенного звонка в игре чемпионата NFC 2019 года. Такая досадная ошибка судейства оказала как внешнее, так и внутреннее давление на лигу, чтобы попытаться избежать подобных вопиющих пропущенных вызовов в будущем, и простым (хотя и ошибочным) решением было сделать фолы подлежащими пересмотру.
Некоторые говорят, что в 2019 году НФЛ не увенчалась успехом в проверке воспроизведения повторов передач с помехами, потому что с самого начала была обречена из-за ошибочной логики. Другие говорят, что эксперимент мог бы сработать, если бы судейский отдел НФЛ решил научиться эффективно использовать проверку PI и не отказывался от этого процесса в течение года.
Несмотря на это, помехи при пасе больше не входят в число розыгрышей/фолов, подпадающих под действие системы просмотра повторов НФЛ. Ниже приведены причины почему.
Можете ли вы оспорить пропуск прохода в НФЛ?
Нет, больше нет. Запросы и невызовы вмешательства в наступательных и защитных пасах подпадали под действие системы проверки повторов НФЛ только в течение одного сезона (2019 г. ). По сути, лига дала этому дополнению к своей сводке правил годовую пробную версию, и вопрос о том, станет ли оно постоянным правилом, будет решаться в 2020 году.
Когда НФЛ пришло время решить, оставить ли вмешательство в пас в качестве подлежащего пересмотру решения, ответом было решительное нет. Фактически, продление проверки PI даже не рассматривалось как вариант.
«Никто снова не выдвигает обзор OPI / DPI, так что это умирает естественной смертью», — сказал член комитета по соревнованиям НФЛ и президент Falcons Рич Маккей на радио SiriusXM NFL во время весеннего собрания владельцев НФЛ.
Отсутствие голосования означало, что поправка к своду правил НФЛ, позволяющая пересматривать вмешательство в пас, будет просто удалена.Что неудивительно, учитывая, что годичный эксперимент был в значительной степени беспорядочным.
«По моему мнению, мы пытались применить то, чего всегда боялись, — сказал Маккей. «Мы не знали, каков будет общий результат, но мы всегда боялись включить в повтор полностью субъективную игру. Большинство повторов объективны, и я имею в виду, что это линия». Пересек самолет или нет? Коснулся ли мяч земли или нет? Был ли он в двух футах или нет? Здесь задействовано много объективных линий, и это объективный обзор.
«Когда мы переходим к субъективному, то есть: «Было ли этого достаточно для фола? Достаточно ли он его схватил? Было ли достаточно ограничения?» Мы знали, что будут проблемы, потому что ваша субъективность и моя субъективность в отношении той или иной рассматриваемой игры могут легко различаться.То, был ли мяч пойман или не пойман — мы можем не согласиться один раз из 100 (раз), но примерно в 99 процентах случаев.
«Это неправда в субъективном мире, поэтому я думаю, когда вы сделали то, что сделали мы, то есть попытались поймать эту действительно вопиющую грандиозную игру и поставить этот стандарт ясности и очевидности, даже тогда, я думаю, вы добавляете субъективный стандарт к субъективной игре, и я думаю, что мы поставили перед собой задачу не прийти к соглашению о результатах, и я думаю, что это проявилось в течение года. »
В дополнение к проблемам субъективности, проверка НФЛ, отказавшаяся от PI, подтвердила, что лига годом ранее просто слишком остро отреагировала на отсутствие вызова, который стоил Святым поездки на Суперкубок 53.
Мы не можем винить НФЛ за Принятие мер после подачи углового «Рэмс» Никелла Роби-Коулмана, блистательного ресивера «Святых» Томмили Льюиса, не вызвали из-за очевидной помехи пасу, учитывая, что судейская ошибка произошла в такой решающий момент в игре чемпионата NFC.Но, как мы писали здесь ранее , лига решила не ту проблему.
Проблема была не в том, что такой вопиющий пропущенный звонок нельзя было просмотреть. Проблема была в том, что такой простой звонок был пропущен.
Майк ДеКурси из SN считает, что эксперимент с проверкой PI мог бы сработать, если бы отдел судейства НФЛ подошёл к процессу более добросовестно.
«В течение сезона 2019 года тренеры лиги оспаривали вызовы о помехах при передаче 81 раз», — написал ДеКурси в мае, когда правило было отменено. «Они выиграли эти испытания 13 раз.Это 16 процентов успеха. Это может показаться мечтательным для Майка Томлина из «Стилерс», который провел сезоны 2017 и 18 годов, не справившись ни с одной задачей, но средний показатель лиги по всем обзорам повторов за 2019 год составил 47 процентов.
«Тренеры добились столь незначительного успеха в первой половине прошлого сезона — всего пять разворотов из 56 попыток вызова за девять недель — что они разочаровались даже в том, чтобы подумать о том, чтобы потратить один из двух выделенных им повторных вызовов на споры о помехах при передаче.
«Трудно интерпретировать это как что-либо иное, кроме как диктовать политику в штаб-квартире… «Это уходит, потому что определенные люди, очевидно, хотели, чтобы это исчезло. Он потерпел неудачу не сам по себе, а как продукт бескорыстного исполнения. Эксперимент не с треском провалился. Экспериментаторы так и сделали.»
Правила помех при пасах НФЛ
Даже в тех случаях, когда можно было пересматривать вызовы или отказы от паса в нападении и защите, официальный язык, касающийся фолов в своде правил НФЛ, не изменился. Он остается таким же и в 2020 году.
Штрафы за помехи при пасе защиты по-прежнему даруют нападающей команде первый даун с точки фола, а если PI также является персональным фолом, применяется дополнительный 15-ярдовый штраф. Штрафы за помехи при передаче в нападении по-прежнему стоили команде 10 ярдов от предыдущей точки.
Прямо из свода правил НФЛ, ниже приведены стандарты, которые лига использует для определения помех при передаче.
Действия, представляющие собой помеху при пасе, включают, но не ограничиваются:
Контакт игрока, не играющего в мяч, который ограничивает возможность соперника поймать мяч;
Игра через спину соперника с целью игры с мячом;
Захват руки(-ей) соперника таким образом, что ограничивает его возможность поймать пас;
Протягивание руки через тело соперника, что ограничивает его возможность поймать передачу, и независимо от того, играет ли игрок, совершающий такое действие, с мячом;
Отрезание пути соперника путем контакта с ним, без игры в мяч;
Захват соперника хуком в попытке добраться до мяча таким образом, что он заставляет тело соперника повернуться до прибытия мяча; или
Начало контакта с противником путем толкания или отталкивания, что создает разделение.
В своде правил НФЛ также перечислены «разрешенные действия обеих команд, когда мяч находится в воздухе».
Другими словами, нижеследующее является честной игрой и не считается вмешательством в пас.
Случайный контакт руками, руками или телом соперника, когда оба игрока борются за мяч или ни один из игроков не ищет мяч. Если есть какие-либо сомнения в том, что контакт был случайным, решение не должно вмешиваться.
Непреднамеренное спутывание стоп, когда оба игрока играют в мяч или ни один из игроков не играет в мяч.
Контакт, который обычно считается помехой при передаче, но передача явно не может быть перехвачена вовлеченными игроками, за исключением случаев, связанных с блокировкой поля нападением.
Возложение руки на соперника, которое не ограничивает его в попытке сыграть с мячом.
Контакт игрока, занявшего позицию над соперником в попытке поймать мяч.



 … Seungminmua11
… Seungminmua11 baby 11:58 Ну вы и долбаёбы) Bfl.baby 11:59 Продолжайте, я уже взял попкорн Jeong-in_F-k 11:59 А я курочку Bfl.baby 12:00 Вы 0хуели Sexy_Soul 12:00 Джисооооооооооон Oh_my.Blblblblbl 12:00 Чо SuperSquirrel
baby 11:58 Ну вы и долбаёбы) Bfl.baby 11:59 Продолжайте, я уже взял попкорн Jeong-in_F-k 11:59 А я курочку Bfl.baby 12:00 Вы 0хуели Sexy_Soul 12:00 Джисооооооооооон Oh_my.Blblblblbl 12:00 Чо SuperSquirrel
 Blblblblbl 12:16 .. SuperSquirrel 12:17 Боже, Сан SuperSquirrel 12:17 Всё вот ты, да опошлишь :00
Blblblblbl 12:16 .. SuperSquirrel 12:17 Боже, Сан SuperSquirrel 12:17 Всё вот ты, да опошлишь :00  Blblblblbl
Blblblblbl ~ won9erlan9 16:14
~ won9erlan9 16:14











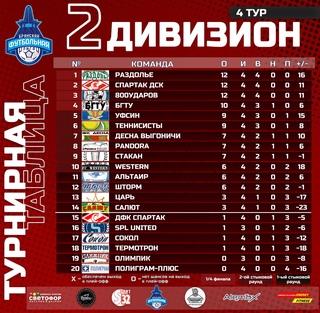
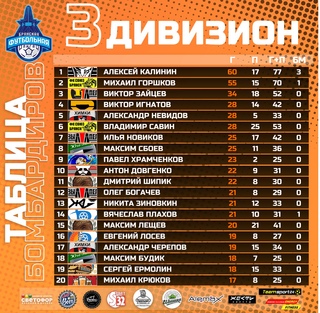

 5GH/s, потребовалось использование двух чипов. А итоговое энергопотребление составило ~6W на 1GH/s.
5GH/s, потребовалось использование двух чипов. А итоговое энергопотребление составило ~6W на 1GH/s. Особенно, если учесть, что это первый ASIC продукт для BFL. Фактические сроки оказались вполне типичными для данной индустрии, ведь, например, у Intel уходит около года с появления первых тестовых образцов и до начала поставок.
Особенно, если учесть, что это первый ASIC продукт для BFL. Фактические сроки оказались вполне типичными для данной индустрии, ведь, например, у Intel уходит около года с появления первых тестовых образцов и до начала поставок.
 д. Набор лицензий на эти средства стоил бы немалых денег где-нибудь в США или Европе, но в Китае и лицензии, как и рабочая сила, оказались гораздо дешевле.
д. Набор лицензий на эти средства стоил бы немалых денег где-нибудь в США или Европе, но в Китае и лицензии, как и рабочая сила, оказались гораздо дешевле.

 Как и ASICMINER, компания расположена в Китае.
Как и ASICMINER, компания расположена в Китае. 5х улучшения производительности, по сравнению с 65нм чипами. История когда BFL пришлось снижать частоту своих чипов из-за ограничений энергопотребления лишь еще раз показывает к чему сейчас приводят попытки вылезти за рамки «дозволенного» тёмным кремнием.
5х улучшения производительности, по сравнению с 65нм чипами. История когда BFL пришлось снижать частоту своих чипов из-за ограничений энергопотребления лишь еще раз показывает к чему сейчас приводят попытки вылезти за рамки «дозволенного» тёмным кремнием.
 Например, использование уровней напряжения, близких к порогу срабатывания (NTV – Near Threshold Voltage). Отсутствие статической памяти и необходимости передавать сигналы на большое расстояние внутри чипа позволяет ожидать здесь еще 5х снижения энергопотребления.
Например, использование уровней напряжения, близких к порогу срабатывания (NTV – Near Threshold Voltage). Отсутствие статической памяти и необходимости передавать сигналы на большое расстояние внутри чипа позволяет ожидать здесь еще 5х снижения энергопотребления. , а количество специализированных чипов падает год от года.
, а количество специализированных чипов падает год от года. Во-вторых, у университетов есть бесплатный доступ к САПР, стоимостью десятки миллионов долларов, лицензированным только для некоммерческого использования. Это означает, что выпускник университета просто не знает как делать «железо» дешево, не разорившись на лицензиях.
Во-вторых, у университетов есть бесплатный доступ к САПР, стоимостью десятки миллионов долларов, лицензированным только для некоммерческого использования. Это означает, что выпускник университета просто не знает как делать «железо» дешево, не разорившись на лицензиях.




 90 007
90 007 2022
2022 Гелинас, CABM и кафедра биохимии, Медицинская школа Роберта Вуда Джонсона, CABM, комната 137, 679 Hoes Lane, Piscataway, NJ 08854-5638, США.ude.sregtur.mbac@sanileg
Гелинас, CABM и кафедра биохимии, Медицинская школа Роберта Вуда Джонсона, CABM, комната 137, 679 Hoes Lane, Piscataway, NJ 08854-5638, США.ude.sregtur.mbac@sanileg Мы показываем, что в живых клетках Bfl-1 избирательно взаимодействует с Bak и tBid, но не с Bax или Bid. Взаимодействие Bfl-1/Bak является функциональным, поскольку Bfl-1 подавляет индуцированный стауроспорином (STS) апоптоз в клетках дикого типа и Вах-дефицитных клетках, но не в клетках Bak-/-. Мы также показываем, что Bfl-1 косвенно блокирует активацию Bax, индуцированную фактором некроза опухоли-α (TNFα), через связь с tBid.Делеция С-конца снижала взаимодействие Bfl-1 с Bak и tBid и снижала его способность подавлять Bak- и tBid-опосредованную гибель клеток. Эти данные показывают, что Bfl-1 использует различные механизмы для подавления апоптоза в зависимости от стимула. Bfl-1 связывается с tBid, чтобы предотвратить активацию проапоптотических Bax и Bak, а также напрямую взаимодействует с Bak, чтобы противодействовать опосредованной Bak гибели клеток, подобно Mcl-1. Таким образом, часть защитной функции NF-κB заключается в индукции Mcl-1-подобной активности путем усиления Bfl-1.
Мы показываем, что в живых клетках Bfl-1 избирательно взаимодействует с Bak и tBid, но не с Bax или Bid. Взаимодействие Bfl-1/Bak является функциональным, поскольку Bfl-1 подавляет индуцированный стауроспорином (STS) апоптоз в клетках дикого типа и Вах-дефицитных клетках, но не в клетках Bak-/-. Мы также показываем, что Bfl-1 косвенно блокирует активацию Bax, индуцированную фактором некроза опухоли-α (TNFα), через связь с tBid.Делеция С-конца снижала взаимодействие Bfl-1 с Bak и tBid и снижала его способность подавлять Bak- и tBid-опосредованную гибель клеток. Эти данные показывают, что Bfl-1 использует различные механизмы для подавления апоптоза в зависимости от стимула. Bfl-1 связывается с tBid, чтобы предотвратить активацию проапоптотических Bax и Bak, а также напрямую взаимодействует с Bak, чтобы противодействовать опосредованной Bak гибели клеток, подобно Mcl-1. Таким образом, часть защитной функции NF-κB заключается в индукции Mcl-1-подобной активности путем усиления Bfl-1. Белки, содержащие только проапоптотический Bh4 (Bid, Bad, Bim, Puma, Noxa, Hrk, Bmf, Nbk/Bik), инициируют передачу сигналов апоптоза в ответ на вызывающие смерть стимулы. Многодоменные белки Bax и Bak являются важными эффекторами апоптоза, и клетки, лишенные обоих белков, невосприимчивы к апоптозу, индуцированному широким спектром стимулов.Напротив, антиапоптотические члены семейства Bcl-2 Bcl-2, Bcl-x L , Bfl-1/A1, Mcl-1 и Bcl-w защищают клетки от апоптоза. Некоторые белки, содержащие только Bh4, такие как tBid и Bim, запускают конформационную активацию Bax и Bak, в то время как другие, такие как Bad, Noxa и Bik, секвестрируют антиапоптотические члены семейства Bcl-2 и нейтрализуют их защитную активность (Letai et al. , 2002). ; Chen и др., 2005; Kuwana и др. , 2005).
Белки, содержащие только проапоптотический Bh4 (Bid, Bad, Bim, Puma, Noxa, Hrk, Bmf, Nbk/Bik), инициируют передачу сигналов апоптоза в ответ на вызывающие смерть стимулы. Многодоменные белки Bax и Bak являются важными эффекторами апоптоза, и клетки, лишенные обоих белков, невосприимчивы к апоптозу, индуцированному широким спектром стимулов.Напротив, антиапоптотические члены семейства Bcl-2 Bcl-2, Bcl-x L , Bfl-1/A1, Mcl-1 и Bcl-w защищают клетки от апоптоза. Некоторые белки, содержащие только Bh4, такие как tBid и Bim, запускают конформационную активацию Bax и Bak, в то время как другие, такие как Bad, Noxa и Bik, секвестрируют антиапоптотические члены семейства Bcl-2 и нейтрализуют их защитную активность (Letai et al. , 2002). ; Chen и др., 2005; Kuwana и др. , 2005). Во внешнем пути лиганды, такие как фактор некроза опухоли-α (TNFα), вызывают опосредованную рецептором смерти активацию каспазы-8, которая расщепляет Bid до его активной формы tBid. В свою очередь, tBid индуцирует конформационную активацию и олигомеризацию Bax и Bak, высвобождение митохондриального цито C , активацию каспазы и апоптоз (Perez and White, 2000; Wei et al., 2000). В то время как антиапоптотические Bcl-2 и Bcl-x L могут связываться как с Bid, так и с tBid, Mcl-1 ингибирует апоптоз, индуцированный рецептором смерти, путем селективного связывания с tBid, но не с Bid (Cheng et al. , 2001; Clohessy). и др. , 2006). Более того, Mcl-1 изолирует эндогенный Bak, но не Bax в здоровых клетках, и блокирует Bak-опосредованную гибель клеток, а Bcl-2 этого не делает (Cuconati et al. , 2003; Willis et al. , 2005). Аденовирусный гомолог Bcl-2 E1B 19K препятствует гибели клеток, связываясь непосредственно с Bax и Bak, но не с tBid или Bid (Oltvai et al.
Во внешнем пути лиганды, такие как фактор некроза опухоли-α (TNFα), вызывают опосредованную рецептором смерти активацию каспазы-8, которая расщепляет Bid до его активной формы tBid. В свою очередь, tBid индуцирует конформационную активацию и олигомеризацию Bax и Bak, высвобождение митохондриального цито C , активацию каспазы и апоптоз (Perez and White, 2000; Wei et al., 2000). В то время как антиапоптотические Bcl-2 и Bcl-x L могут связываться как с Bid, так и с tBid, Mcl-1 ингибирует апоптоз, индуцированный рецептором смерти, путем селективного связывания с tBid, но не с Bid (Cheng et al. , 2001; Clohessy). и др. , 2006). Более того, Mcl-1 изолирует эндогенный Bak, но не Bax в здоровых клетках, и блокирует Bak-опосредованную гибель клеток, а Bcl-2 этого не делает (Cuconati et al. , 2003; Willis et al. , 2005). Аденовирусный гомолог Bcl-2 E1B 19K препятствует гибели клеток, связываясь непосредственно с Bax и Bak, но не с tBid или Bid (Oltvai et al. , 1993; Перес и Уайт, 2000 г.; Куконати и др. , 2002; Cuconati and White, 2002), но как другие антиапоптотические члены семейства Bcl-2, такие как Bfl-1, блокируют апоптоз, не совсем ясно.
, 1993; Перес и Уайт, 2000 г.; Куконати и др. , 2002; Cuconati and White, 2002), но как другие антиапоптотические члены семейства Bcl-2, такие как Bfl-1, блокируют апоптоз, не совсем ясно. Сообщалось, что человеческий Bfl-1 взаимодействует с Bid в временно трансфицированных клетках и с рекомбинантным tBid in vitro , но не с Bax или Bak (Werner et al. , 2002). Было описано, что Bfl-1/A1 связывается с Bax в двухгибридных дрожжах или путем совместной иммунопреципитации in vitro транслированных белков (Sedlak et al. , 1995; Zhang et al. , 2000). Другие исследователи не обнаружили связи между A1 мыши и Bax в котрансфицированных клетках, но A1 взаимодействовал с Bak в этих условиях (Holmgreen et al., 1999). Здесь мы исследовали связь Bfl-1 с эндогенными Bax, Bak, Bid и tBid в живых клетках и изучили, как эти взаимодействия связаны с его защитной активностью в ответ на внешние и внутренние стимулы, вызывающие смерть.
Сообщалось, что человеческий Bfl-1 взаимодействует с Bid в временно трансфицированных клетках и с рекомбинантным tBid in vitro , но не с Bax или Bak (Werner et al. , 2002). Было описано, что Bfl-1/A1 связывается с Bax в двухгибридных дрожжах или путем совместной иммунопреципитации in vitro транслированных белков (Sedlak et al. , 1995; Zhang et al. , 2000). Другие исследователи не обнаружили связи между A1 мыши и Bax в котрансфицированных клетках, но A1 взаимодействовал с Bak в этих условиях (Holmgreen et al., 1999). Здесь мы исследовали связь Bfl-1 с эндогенными Bax, Bak, Bid и tBid в живых клетках и изучили, как эти взаимодействия связаны с его защитной активностью в ответ на внешние и внутренние стимулы, вызывающие смерть. Bfl-1 был помечен с N-конца зеленым флуоресцентным белком (GFP), поскольку коммерческие антитела не смогли успешно распознать Bfl-1 человека. GFP-Bfl-1 локализуется в митохондриях и околоядерной области, перекрывая эндогенный цито C (). В то время как TNFα провоцировал высвобождение цито C в экспрессирующих GFP клетках, цито C оставался митохондриально локализованным в GFP-Bfl-1-положительных клетках, что согласуется с его защитной активностью в отношении TNFα (Karsan et al. , 1996; Wang ). и другие., 1999; Зонг и др. , 1999). Мутант GFP-Bfl-1ΔC, лишенный С-концевых 24 аминокислот, распространялся по всей клетке, включая ядро, подобно GFP, но все еще подавлял TNFα-индуцированное высвобождение цито C . Высвобождение Cyto C и апоптоз критически зависят от конформационной активации и олигомеризации Bax и Bak, которые ингибируются антиапоптотическими Bcl-2 и Bcl-x L . Точно так же GFP-Bfl-1 и Bfl-1ΔC подавляли TNFα-индуцированную конформационную активацию Bax, как видно из антитела, специфичного к N-концу Bax, которое маскируется, когда Bax неактивен, и экспонируется при активации Bax (; Desagher et al.
Bfl-1 был помечен с N-конца зеленым флуоресцентным белком (GFP), поскольку коммерческие антитела не смогли успешно распознать Bfl-1 человека. GFP-Bfl-1 локализуется в митохондриях и околоядерной области, перекрывая эндогенный цито C (). В то время как TNFα провоцировал высвобождение цито C в экспрессирующих GFP клетках, цито C оставался митохондриально локализованным в GFP-Bfl-1-положительных клетках, что согласуется с его защитной активностью в отношении TNFα (Karsan et al. , 1996; Wang ). и другие., 1999; Зонг и др. , 1999). Мутант GFP-Bfl-1ΔC, лишенный С-концевых 24 аминокислот, распространялся по всей клетке, включая ядро, подобно GFP, но все еще подавлял TNFα-индуцированное высвобождение цито C . Высвобождение Cyto C и апоптоз критически зависят от конформационной активации и олигомеризации Bax и Bak, которые ингибируются антиапоптотическими Bcl-2 и Bcl-x L . Точно так же GFP-Bfl-1 и Bfl-1ΔC подавляли TNFα-индуцированную конформационную активацию Bax, как видно из антитела, специфичного к N-концу Bax, которое маскируется, когда Bax неактивен, и экспонируется при активации Bax (; Desagher et al. ., 1999; Перес и Уайт, 2000). Напротив, Bax активировался в окружающих клетках, в которых отсутствовала значительная экспрессия GFP-Bfl-1. Таким образом, Bfl-1 и Bfl-1ΔC подавляют активацию Bax в ответ на TNFα.
., 1999; Перес и Уайт, 2000). Напротив, Bax активировался в окружающих клетках, в которых отсутствовала значительная экспрессия GFP-Bfl-1. Таким образом, Bfl-1 и Bfl-1ΔC подавляют активацию Bax в ответ на TNFα. Клетки также анализировали на флуоресценцию GFP ( b , e , h , l , p , t ). Цито С , цитохром С ; GFP, зеленый флуоресцентный белок; TNFα, фактор некроза опухоли-α.
Клетки также анализировали на флуоресценцию GFP ( b , e , h , l , p , t ). Цито С , цитохром С ; GFP, зеленый флуоресцентный белок; TNFα, фактор некроза опухоли-α. Клетки также анализировали на флуоресценцию GFP (—, -с-, —, —, -1-, —). Звездочками отмечены GFP-Bfl-1 или -Bfl-1 AC-положительные клетки, которые не показали активации Bax.
Клетки также анализировали на флуоресценцию GFP (—, -с-, —, —, -1-, —). Звездочками отмечены GFP-Bfl-1 или -Bfl-1 AC-положительные клетки, которые не показали активации Bax. , 1998). Хотя Bfl-1 и Bfl-1ΔC ко-иммунопреципитируются со сверхэкспрессированным FLAG-Bax (1), ни один из них не связан ни с эндогенным Bax, активированным TNFα/циклогексимидом (CHX), ни с неактивным Bax в клетках, обработанных CHX, несмотря на эффективную иммунопреципитацию Bax (2).Обратная иммунопреципитация эндогенного Bax с помощью GFP-Bfl-1 или 2×Myc-Bfl-1 также не удалась, в отличие от контрольной иммунопреципитации эндогенного Bax с помощью Bak (14). Т.о., хотя Bfl-1 может ассоциироваться со сверхэкспрессией Bax, комплексов с эндогенным Bax не наблюдалось, что свидетельствует о том, что он противодействует активации Bax, взаимодействуя с факторами, действующими выше каскада TNF α.
, 1998). Хотя Bfl-1 и Bfl-1ΔC ко-иммунопреципитируются со сверхэкспрессированным FLAG-Bax (1), ни один из них не связан ни с эндогенным Bax, активированным TNFα/циклогексимидом (CHX), ни с неактивным Bax в клетках, обработанных CHX, несмотря на эффективную иммунопреципитацию Bax (2).Обратная иммунопреципитация эндогенного Bax с помощью GFP-Bfl-1 или 2×Myc-Bfl-1 также не удалась, в отличие от контрольной иммунопреципитации эндогенного Bax с помощью Bak (14). Т.о., хотя Bfl-1 может ассоциироваться со сверхэкспрессией Bax, комплексов с эндогенным Bax не наблюдалось, что свидетельствует о том, что он противодействует активации Bax, взаимодействуя с факторами, действующими выше каскада TNF α. ( b ) Иммунопреципитация 2×Myc-Bfl-1 или -Bfl-1ΔC с эндогенным Bax в клетках HeLa, обработанных TNFα плюс CHX или только CHX, с использованием антител против Bax-loop и иммуноблоттинга с анти-Myc или -BaxN20 антитело. Lys: 1/10 вводного лизата. CHX, циклогексимид; GFP, зеленый флуоресцентный белок; TNFα, фактор некроза опухоли-α.
( b ) Иммунопреципитация 2×Myc-Bfl-1 или -Bfl-1ΔC с эндогенным Bax в клетках HeLa, обработанных TNFα плюс CHX или только CHX, с использованием антител против Bax-loop и иммуноблоттинга с анти-Myc или -BaxN20 антитело. Lys: 1/10 вводного лизата. CHX, циклогексимид; GFP, зеленый флуоресцентный белок; TNFα, фактор некроза опухоли-α. ( d ) Иммортализованные iBMK дикого типа, Bak-/-, Bax-/- и Bax-/-Bak-/- с двойным нокаутом, трансфицированные EGFP (заштрихованы), 2×Myc-Bfl-1 (черные), 2×Myc-Bfl-1ΔC (белый) или FLAG-Mcl-1 (точечный) обрабатывали STS в течение 24 ч или оставляли без обработки. Среднюю выживаемость в трех экспериментах определяли по исключению трипанового синего и нормализовали по эффективности трансфекции. CHX, циклогексимид; GFP, зеленый флуоресцентный белок; iBMK, бессмертная почка мышиного детеныша; STS, стауроспорин; TNFα, фактор некроза опухоли-α.
( d ) Иммортализованные iBMK дикого типа, Bak-/-, Bax-/- и Bax-/-Bak-/- с двойным нокаутом, трансфицированные EGFP (заштрихованы), 2×Myc-Bfl-1 (черные), 2×Myc-Bfl-1ΔC (белый) или FLAG-Mcl-1 (точечный) обрабатывали STS в течение 24 ч или оставляли без обработки. Среднюю выживаемость в трех экспериментах определяли по исключению трипанового синего и нормализовали по эффективности трансфекции. CHX, циклогексимид; GFP, зеленый флуоресцентный белок; iBMK, бессмертная почка мышиного детеныша; STS, стауроспорин; TNFα, фактор некроза опухоли-α. Взаимодействие Bfl-1ΔC с Bak было значительно слабее, чем у Bfl-1 ().Таким образом, Bfl-1 ассоциирует с неактивной и активной формами эндогенного Bak, тогда как сниженное взаимодействие Bfl-1ΔC, скорее всего, является результатом его делокализации из митохондрий.
Взаимодействие Bfl-1ΔC с Bak было значительно слабее, чем у Bfl-1 ().Таким образом, Bfl-1 ассоциирует с неактивной и активной формами эндогенного Bak, тогда как сниженное взаимодействие Bfl-1ΔC, скорее всего, является результатом его делокализации из митохондрий. Как и в случае с Bfl-1, Bfl-1ΔC не способен подавлять апоптоз в клетках Bak-/-. Это показало, что селективное взаимодействие Bfl-1 с Bak, но не с Bax, коррелирует с подавлением Bak-, но не Bax-опосредованного апоптоза, подобно Mcl-1.
Как и в случае с Bfl-1, Bfl-1ΔC не способен подавлять апоптоз в клетках Bak-/-. Это показало, что селективное взаимодействие Bfl-1 с Bak, но не с Bax, коррелирует с подавлением Bak-, но не Bax-опосредованного апоптоза, подобно Mcl-1. , 2005; Herold et al. , 2006). Результаты заметно отличались, когда мы исследовали ассоциацию Bfl-1 с эндогенным tBid в клетках, обработанных TNFα/CHX, чтобы способствовать процессингу Bid в tBid (). Хотя эндогенный tBid трудно обнаружить из-за его короткого периода полувыведения (Breitschopf et al., 2000), было ясно видно взаимодействие с 2×Myc-Bfl-1 или Bfl-1ΔC, особенно при более длительном воздействии. В поддержку функционального взаимодействия Bfl-1 и Bfl-1ΔC подавляли апоптоз, индуцированный tBid, хотя Bfl-1ΔC был несколько менее эффективен, чем Bfl-1 (12). Это показывает, что подобно его дифференциальному взаимодействию с эндогенным Bak, но не с Bax, Bfl-1 избирательно связывается с эндогенным tBid, но не с Bid, и предполагает, что это взаимодействие важно для подавления внешнего апоптотического каскада, инициируемого TNFα.
, 2005; Herold et al. , 2006). Результаты заметно отличались, когда мы исследовали ассоциацию Bfl-1 с эндогенным tBid в клетках, обработанных TNFα/CHX, чтобы способствовать процессингу Bid в tBid (). Хотя эндогенный tBid трудно обнаружить из-за его короткого периода полувыведения (Breitschopf et al., 2000), было ясно видно взаимодействие с 2×Myc-Bfl-1 или Bfl-1ΔC, особенно при более длительном воздействии. В поддержку функционального взаимодействия Bfl-1 и Bfl-1ΔC подавляли апоптоз, индуцированный tBid, хотя Bfl-1ΔC был несколько менее эффективен, чем Bfl-1 (12). Это показывает, что подобно его дифференциальному взаимодействию с эндогенным Bak, но не с Bax, Bfl-1 избирательно связывается с эндогенным tBid, но не с Bid, и предполагает, что это взаимодействие важно для подавления внешнего апоптотического каскада, инициируемого TNFα. ( a ) Иммунопреципитация/вестерн-блоттинг GFP-Bfl-1 или -Bfl-1ΔC со сверхэкспрессией Bid-Myc или tBid-Myc в клетках HeLa. ( b ) Коиммунопреципитация эндогенного Bid или tBid с 2×Myc-Bfl-1 или -Bfl-1ΔC в клетках HeLa, обработанных только TNFα/CHX или CHX. Показана более длительная экспозиция. Звездочками отмечены позиции эндогенных Bid и tBid; над Bid в лизатах появлялась неспецифическая полоса.( c ) GFP-Bfl-1 или -Bfl-1ΔC подавляют вызванную tBid гибель клеток в клетках MCF-7, котрансфицированных tBid-FLAG или pcDNA3 вместе с CMV-β-gal. Показано среднее из трех экспериментов. ( d ) Модель, иллюстрирующая избирательное взаимодействие Bfl-1 с tBid и Bak для подавления TNFα- и Bak-зависимого апоптоза, а также сходство его механизма действия с Mcl-1. CHX, циклогексимид; GFP, зеленый флуоресцентный белок; TNFα, фактор некроза опухоли-α.
( a ) Иммунопреципитация/вестерн-блоттинг GFP-Bfl-1 или -Bfl-1ΔC со сверхэкспрессией Bid-Myc или tBid-Myc в клетках HeLa. ( b ) Коиммунопреципитация эндогенного Bid или tBid с 2×Myc-Bfl-1 или -Bfl-1ΔC в клетках HeLa, обработанных только TNFα/CHX или CHX. Показана более длительная экспозиция. Звездочками отмечены позиции эндогенных Bid и tBid; над Bid в лизатах появлялась неспецифическая полоса.( c ) GFP-Bfl-1 или -Bfl-1ΔC подавляют вызванную tBid гибель клеток в клетках MCF-7, котрансфицированных tBid-FLAG или pcDNA3 вместе с CMV-β-gal. Показано среднее из трех экспериментов. ( d ) Модель, иллюстрирующая избирательное взаимодействие Bfl-1 с tBid и Bak для подавления TNFα- и Bak-зависимого апоптоза, а также сходство его механизма действия с Mcl-1. CHX, циклогексимид; GFP, зеленый флуоресцентный белок; TNFα, фактор некроза опухоли-α. Мы демонстрируем, что в функциональном контексте живых клеток, где Bfl-1 ингибирует апоптоз, Bfl-1 избирательно связывается с эндогенными tBid и Bak, но не с Bid или Bax. В то время как Bfl-1 противодействовал активации Bax в ответ на внешнюю передачу сигнала о смерти с помощью TNFα, он не смог подавить Bax-опосредованную гибель клеток в ответ на внутреннюю передачу сигнала о смерти, которая не зависит от tBid в клетках Bak-/-, но эффективно блокировал STS-индуцированный апоптоз. в клетках Bax-/-, которые экспрессируют эндогенный Bak. Это указывает на то, что конститутивное взаимодействие Bfl-1 с Bak важно для подавления Bak-опосредованного апоптоза, тогда как его связь с tBid, скорее всего, ответственна за блокирование активации Bax в ответ на стимуляцию рецептора смерти.
Мы демонстрируем, что в функциональном контексте живых клеток, где Bfl-1 ингибирует апоптоз, Bfl-1 избирательно связывается с эндогенными tBid и Bak, но не с Bid или Bax. В то время как Bfl-1 противодействовал активации Bax в ответ на внешнюю передачу сигнала о смерти с помощью TNFα, он не смог подавить Bax-опосредованную гибель клеток в ответ на внутреннюю передачу сигнала о смерти, которая не зависит от tBid в клетках Bak-/-, но эффективно блокировал STS-индуцированный апоптоз. в клетках Bax-/-, которые экспрессируют эндогенный Bak. Это указывает на то, что конститутивное взаимодействие Bfl-1 с Bak важно для подавления Bak-опосредованного апоптоза, тогда как его связь с tBid, скорее всего, ответственна за блокирование активации Bax в ответ на стимуляцию рецептора смерти. , 2002; Grinberg et al. , 2002). Однако исследования нокаута показали, что tBid, скорее всего, действует путем активации Bax и Bak, поскольку клетки, лишенные обоих факторов, устойчивы к апоптозу (Wei et al. , 2001; Degenhardt et al., 2002). Наши данные подтверждают идею о том, что ассоциация Bfl-1 с tBid, скорее всего, ответственна за подавление активации Bax, поскольку Bfl-1 не смог связать эндогенный Bax в клетках, обработанных TNF. Это согласуется с сообщениями, показывающими, что, хотя Bcl-2 и Bcl-x L могут напрямую ассоциироваться с Bax, их взаимодействие с tBid наиболее важно для их защитной функции в каскаде TNF (Cheng et al. , 2001; Werner ). и др. , 2002; Yi и др. , 2003).
, 2002; Grinberg et al. , 2002). Однако исследования нокаута показали, что tBid, скорее всего, действует путем активации Bax и Bak, поскольку клетки, лишенные обоих факторов, устойчивы к апоптозу (Wei et al. , 2001; Degenhardt et al., 2002). Наши данные подтверждают идею о том, что ассоциация Bfl-1 с tBid, скорее всего, ответственна за подавление активации Bax, поскольку Bfl-1 не смог связать эндогенный Bax в клетках, обработанных TNF. Это согласуется с сообщениями, показывающими, что, хотя Bcl-2 и Bcl-x L могут напрямую ассоциироваться с Bax, их взаимодействие с tBid наиболее важно для их защитной функции в каскаде TNF (Cheng et al. , 2001; Werner ). и др. , 2002; Yi и др. , 2003). , 2005), хотя некоторые обнаружили иное (Certo et al. , 2006).Наш вывод о том, что Bfl-1 физически и функционально взаимодействует с Bak в живых клетках, отличается от других, которым не удалось увидеть взаимодействие между , транслируемым in vitro Bfl-1, и Bak в присутствии митохондрий печени мыши (Werner et al. , 2002). ). Поскольку рекомбинантный tBid был включен в эти анализы, а ассоциация tBid с Mcl-1 может вытеснять Bak из комплексов Mcl-1-Bak (Clohessy et al. , 2006), tBid также может диссоциировать Bfl-1 от Bak, тем самым препятствуя обнаружению комплексов Bfl-1-Bak в этих анализах.
, 2005), хотя некоторые обнаружили иное (Certo et al. , 2006).Наш вывод о том, что Bfl-1 физически и функционально взаимодействует с Bak в живых клетках, отличается от других, которым не удалось увидеть взаимодействие между , транслируемым in vitro Bfl-1, и Bak в присутствии митохондрий печени мыши (Werner et al. , 2002). ). Поскольку рекомбинантный tBid был включен в эти анализы, а ассоциация tBid с Mcl-1 может вытеснять Bak из комплексов Mcl-1-Bak (Clohessy et al. , 2006), tBid также может диссоциировать Bfl-1 от Bak, тем самым препятствуя обнаружению комплексов Bfl-1-Bak в этих анализах. Это может объяснить, почему в недавней работе с использованием Bfl-1ΔC-подобной формы A1, лишенной 20 C-концевых аминокислот, не удалось обнаружить взаимодействие с Bak (Chen et al. , 2005; Willis et al. , 2005). ). Ослабленная ассоциация Bfl-1ΔC с Bak может также прояснить, почему делеция C-конца Bfl-1 снижает его защитную активность в отношении p53-индуцированного апоптоза, в котором Bak играет важную роль (D’Sa-Eipper and Chinnadurai, 1998). В совокупности эти находки показывают, что эффективное взаимодействие Bfl-1/Bak может быть важным средством для Bfl-1 противодействовать внутренней передаче сигналов смерти, которая зависит от Bak и не передает сигналы через tBid.
Это может объяснить, почему в недавней работе с использованием Bfl-1ΔC-подобной формы A1, лишенной 20 C-концевых аминокислот, не удалось обнаружить взаимодействие с Bak (Chen et al. , 2005; Willis et al. , 2005). ). Ослабленная ассоциация Bfl-1ΔC с Bak может также прояснить, почему делеция C-конца Bfl-1 снижает его защитную активность в отношении p53-индуцированного апоптоза, в котором Bak играет важную роль (D’Sa-Eipper and Chinnadurai, 1998). В совокупности эти находки показывают, что эффективное взаимодействие Bfl-1/Bak может быть важным средством для Bfl-1 противодействовать внутренней передаче сигналов смерти, которая зависит от Bak и не передает сигналы через tBid. , 2002, 2003; Clohessy и др. , 2006). Хотя Bfl-1 и Mcl-1 используют сходные средства для подавления апоптоза, они, вероятно, действуют неповторяющимся образом в разных клетках и в ответ на разные стимулы, поскольку они находятся под разным транскрипционным контролем, а Mcl-1 не является Мишень NF-κB (Michels et al., 2005). Bfl-1 сверхэкспрессируется во многих опухолях человека, в том числе в некоторых, в которых конститутивная активация NF-κB необходима для выживания и химиорезистентности. Таким образом, функциональное взаимодействие Bfl-1 с tBid и Bak может способствовать его роли в онкогенезе и резистентности к терапии. В совокупности эти исследования позволяют по-новому взглянуть на механизмы, с помощью которых Bfl-1 противодействует внешним и внутренним каскадам сигналов смерти, и позволяют предположить, что подходы к специфической блокировке взаимодействия Bfl-1 с tBid и/или Bak могут улучшить реакцию клеток, экспрессирующих Bfl-1. опухолевых клеток к противораковому лечению.
, 2002, 2003; Clohessy и др. , 2006). Хотя Bfl-1 и Mcl-1 используют сходные средства для подавления апоптоза, они, вероятно, действуют неповторяющимся образом в разных клетках и в ответ на разные стимулы, поскольку они находятся под разным транскрипционным контролем, а Mcl-1 не является Мишень NF-κB (Michels et al., 2005). Bfl-1 сверхэкспрессируется во многих опухолях человека, в том числе в некоторых, в которых конститутивная активация NF-κB необходима для выживания и химиорезистентности. Таким образом, функциональное взаимодействие Bfl-1 с tBid и Bak может способствовать его роли в онкогенезе и резистентности к терапии. В совокупности эти исследования позволяют по-новому взглянуть на механизмы, с помощью которых Bfl-1 противодействует внешним и внутренним каскадам сигналов смерти, и позволяют предположить, что подходы к специфической блокировке взаимодействия Bfl-1 с tBid и/или Bak могут улучшить реакцию клеток, экспрессирующих Bfl-1. опухолевых клеток к противораковому лечению. Это может быть важно, поскольку ингибирование Mcl-1 необходимо для сенсибилизации некоторых опухолей к низкомолекулярным миметикам Bh4 ABT-737, которые противодействуют Bcl-x L , Bcl-2 и Bcl-w (Konopleva et al. ). , 2006; Letai, 2006; van Delft и др. , 2006). Если Bfl-1 представляет собой NF-κB-индуцированный заменитель Mcl-1, антагонистическая функция Bfl-1 может быть необходима для терапии рака и, возможно, является одним из средств, с помощью которых ингибиторы NF-κB провоцируют апоптоз раковых клеток.
Это может быть важно, поскольку ингибирование Mcl-1 необходимо для сенсибилизации некоторых опухолей к низкомолекулярным миметикам Bh4 ABT-737, которые противодействуют Bcl-x L , Bcl-2 и Bcl-w (Konopleva et al. ). , 2006; Letai, 2006; van Delft и др. , 2006). Если Bfl-1 представляет собой NF-κB-индуцированный заменитель Mcl-1, антагонистическая функция Bfl-1 может быть необходима для терапии рака и, возможно, является одним из средств, с помощью которых ингибиторы NF-κB провоцируют апоптоз раковых клеток.
 Дж. Биол. Хим. 2000; 275:21648–21652. [PubMed] [Google Scholar]
Дж. Биол. Хим. 2000; 275:21648–21652. [PubMed] [Google Scholar] Дж. Биол. Хим. 2006; 281:5750–5759. [PubMed] [Google Scholar]
Дж. Биол. Хим. 2006; 281:5750–5759. [PubMed] [Google Scholar] Онкоген. 1998;16:3105–3114. [PubMed] [Google Scholar]
Онкоген. 1998;16:3105–3114. [PubMed] [Google Scholar] , Сариг Р., Зальцман Ю., Фрумкин Д., Грамматикакис Н., Реувени Э. и др. tBID гомоолигомеризуется в митохондриальной мембране, вызывая апоптоз. Дж. Биол. Хим. 2002; 277:12237–12245. [PubMed] [Google Scholar]
, Сариг Р., Зальцман Ю., Фрумкин Д., Грамматикакис Н., Реувени Э. и др. tBID гомоолигомеризуется в митохондриальной мембране, вызывая апоптоз. Дж. Биол. Хим. 2002; 277:12237–12245. [PubMed] [Google Scholar] Дж. Биол. Хим. 1998; 273:10777–10783. [PubMed] [Google Scholar]
Дж. Биол. Хим. 1998; 273:10777–10783. [PubMed] [Google Scholar] Мол Ячейка. 2005; 17: 525–535. [PubMed] [Google Scholar]
Мол Ячейка. 2005; 17: 525–535. [PubMed] [Google Scholar] Дж. Биол. Хим. 1993; 268:25265–25268. [PubMed] [Google Scholar]
Дж. Биол. Хим. 1993; 268:25265–25268. [PubMed] [Google Scholar] Миметик Bh4 ABT-737 нацеливается на селективные белки Bcl-2 и эффективно индуцирует апоптоз через Bak/Bax, если Mcl-1 нейтрализуется.Раковая клетка. 2006; 10: 389–399. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Миметик Bh4 ABT-737 нацеливается на селективные белки Bcl-2 и эффективно индуцирует апоптоз через Bak/Bax, если Mcl-1 нейтрализуется.Раковая клетка. 2006; 10: 389–399. [Бесплатная статья PMC] [PubMed] [Google Scholar] Член семьи Bcl-2 секвестрирует Bfl-1/A1 укорочено, чтобы предотвратить его сотрудничество с проапоптотическим Bak или Бакс.Дж. Биол. Хим. 2002; 277: 22781–22788. [PubMed] [Google Scholar]
Член семьи Bcl-2 секвестрирует Bfl-1/A1 укорочено, чтобы предотвратить его сотрудничество с проапоптотическим Bak или Бакс.Дж. Биол. Хим. 2002; 277: 22781–22788. [PubMed] [Google Scholar] Гомолог Bcl-2 для выживания Bfl-1/A1 является прямой мишенью транскрипции NF-κB, который блокирует апоптоз, индуцированный TNFα. . Гены Дев. 1999; 13: 382–387. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Гомолог Bcl-2 для выживания Bfl-1/A1 является прямой мишенью транскрипции NF-κB, который блокирует апоптоз, индуцированный TNFα. . Гены Дев. 1999; 13: 382–387. [Бесплатная статья PMC] [PubMed] [Google Scholar] Такая досадная ошибка судейства оказала как внешнее, так и внутреннее давление на лигу, чтобы попытаться избежать подобных вопиющих пропущенных вызовов в будущем, и простым (хотя и ошибочным) решением было сделать фолы подлежащими пересмотру.
Такая досадная ошибка судейства оказала как внешнее, так и внутреннее давление на лигу, чтобы попытаться избежать подобных вопиющих пропущенных вызовов в будущем, и простым (хотя и ошибочным) решением было сделать фолы подлежащими пересмотру. ). По сути, лига дала этому дополнению к своей сводке правил годовую пробную версию, и вопрос о том, станет ли оно постоянным правилом, будет решаться в 2020 году.
). По сути, лига дала этому дополнению к своей сводке правил годовую пробную версию, и вопрос о том, станет ли оно постоянным правилом, будет решаться в 2020 году. Большинство повторов объективны, и я имею в виду, что это линия». Пересек самолет или нет? Коснулся ли мяч земли или нет? Был ли он в двух футах или нет? Здесь задействовано много объективных линий, и это объективный обзор.
Большинство повторов объективны, и я имею в виду, что это линия». Пересек самолет или нет? Коснулся ли мяч земли или нет? Был ли он в двух футах или нет? Здесь задействовано много объективных линий, и это объективный обзор. »
» «Они выиграли эти испытания 13 раз.Это 16 процентов успеха. Это может показаться мечтательным для Майка Томлина из «Стилерс», который провел сезоны 2017 и 18 годов, не справившись ни с одной задачей, но средний показатель лиги по всем обзорам повторов за 2019 год составил 47 процентов.
«Они выиграли эти испытания 13 раз.Это 16 процентов успеха. Это может показаться мечтательным для Майка Томлина из «Стилерс», который провел сезоны 2017 и 18 годов, не справившись ни с одной задачей, но средний показатель лиги по всем обзорам повторов за 2019 год составил 47 процентов. Он остается таким же и в 2020 году.
Он остается таким же и в 2020 году.
